|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| Thread ▼ Details |
|
Thread Info
|
|
|
| Author | Topic: An ID hypothesis: Front-loaded Evolution | |||||||||||||||||||||||||
|
RAZD Member (Idle past 1424 days)  Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Genomics, thanks.
I really think that has more to do with our differing perspectives and backgrounds. I don't have that much interest in the origin of the universe, and the origin of natural laws. I am more interested in the history of life on earth. To you, it might seem that FLE is secondary, but to me it does not seem secondary at all. Fair enough. Admittedly I am interested in "the ultimate question of life, the universe, and oh, everything" to quote Douglas Adams, however I am also interested in explaining life on earth as part of that. Perhaps the difference is in focus angles (wide vs narrow).
That's assuming life arose on earth. But this assumes life developed here, on our planet, instead of being delivered to our planet through panspermia. While you assume it arose elsewhere and then was delivered here. We do know that at the early stages of earths formation there was no life that left any traces that we have found, and we do know that the oldest rocks known that could show fossil records of life do so - evidence for cyanobacteria - at some 3.5 billion years ago. A google search on cyanobacteria oldest life brings up a lot of hits: History of life - WikipediaNicht gefunden - Not found For example. What we don't know is how that life got there. Your delivery system requires a previously developed life\being while mine only requires observed systems for delivery of observed materials from space to the surface of a planet. With no evidence of your delivery system you must admit that this is an extremely weak link in your hypothesis: you have to assume that it existed.
Well, this observation can be interpreted differently. Clouds of pre-biotic molecules can easily be the remnants of once-alive cells. ... You forget the age of these interstellar clouds - because of their distance from earth what we are seeing were there before the earth formed. Thus the absence of identifiable interstellar cells is problematic for a panspermia concept.
... Meteorites with nucleic and amino acids could easily be evidence that cells were once present on these meteorites. We wouldn't expect to find large amounts of clouds with bacteria cells, because it's far easier for the chemical constituents of the cells to survive in space than for the cells themselves to survive. Just saying. Now you are assuming that meteorites are your delivery system and that life arose on meteorites? Just saying.
I am not in any way whatsoever proposing that the earth is the result of some special creation. Nor am I proposing that life on earth is the result of "special creation," unless by that you mean engineering, and nothing supernatural. I'm not necessarily saying that life itself is designed, either ,,, So you would agree then that there is nothing special about life existing on earth?
... I am proposing that the initial genomes on earth were engineered to bias evolution. ... While I am proposing that the initial pre-bionic molecules were made to act according to the designed laws of chemistry and evolution, to bias first the formation of life from the provided building blocks, and second what life evolved from the first cells. You say cells (origin unknown), while I say cells with some assembly required (together with instructions, assembly materials and some self-assembly systems).
... We've been able to implement our own designs into already-existing genomes for some time now, but we're still a ways from engineering life itself. And yet you are still only copying an existing system for making life. Building one that uses pre-biotic materials would confirm\validate my position that life was built from pre-biotic materials, would you not agree? Especially when you consider the work being done on self-assembly molecules and pre-cellular life systems. Being able to construct life from pre-biotic materials would be a prediction of FLU but not of FLE, yes? Does FLE predict that this will not be possible? It certainly seems to imply it to me, otherwise why is FLE necessary?
I wouldn't say FLE is an attempt at being a scientific hypothesis. It is one, by the standard definition of a scientific hypothesis, IMHO. It's not a philosophy, like deism is. It's not a faith either, unless you consider hypotheses like flagellar --> TTSS evolution a faith or philosophy. ... type III secretion system (TTSS). ... A scientific hypothesis, as opposed to a philosophical one, is based on objective evidence or objective observation (rather than on personal intuition: see Percy Message 210), AND it is testable. Objective evidence: pre-biotic molecules exist in space, both in distant interstellar clouds and on near earth asteroids, and they have been recovered from meteors impacting earth. Objective evidence: the earth formed 4.5 billion years ago, with lots of meteors impacting the surface in the early years, and life is known to exist 3.5 billion years ago, with no earlier record currently available for either life or precursors to life. Objective observation: the first life on earth occurred between 3.5 billion years ago and 4.5 billion years ago. Scientific hypothesis: life arose between 4.5 billion and 3.5 billion years ago from pre-biotic molecules via "natural (FLU or NFLU) laws" for chemistry and chemical evolutionary processes. Test: replicate how life could have formed to see if it can form as hypothesized. Faith: some unknown being made and delivered a designed front-loaded life system to earth. Test: believe it.
I'm not defending or attacking NFLU or FLU ... I really wouldn't care either way, unless you're going to claim that FLU and FLE are incompatible. Curiously, I'm pointing out that your hypothesis is not testable - or at least you have not demonstrated how it could be properly tested.
... (geez, I almost spelled that "flue"; no offense - humor people, humor ). ... There's also the joke about a flea and a fly in a flu were imprisoned, so what could they do; said the fly let us flee said the flee let us fly so the flew through a flaw in the flu.
Note that I'm not in any way specifying the intelligence behind front-loaded evolution. ... But you have to admit that it is not a scientific hypothesis, but an assumption based on personal intuition\opinion\wish\hope, and that your FLE hypothesis is founded on that assumption. You might as well assume that god/s exist and that god-did-it to provide the first cellular life on earth, yes?
... Having said that, you can test FLE by verifying or disproving its predictions. Confirmation of a prediction of the FLE model is evidence in favor of the FLE model. ... If the prediction is for something that could not occur under FLU or NFLU, then confirmation of a prediction would be evidence in favor of it. If the prediction is for something that cannot differentiate FLE from FLU or NFLU, then confirmation of a prediction just means that it is not falsified, while the validity of the hypothesis is not tested. You should also have a falsification test for something that could occur via FLU/NFLU natural laws but would not occur under FLE -- do you have such a test?
... This does not mean you can rule out the possibility that purely non-telic mechanisms can account for x-y-z. But if non-telic mechanisms do not predict x-y-z, while FLE does, then it is evidence for FLE. This is how science works. The FLU model does not predict that deep homology prediction made by the FLE model; nor does conventional theory predict it. Both the FLU and non-telic models can account for that observation, but they do not predict it, while the FLE model does, so confirmation of that prediction is evidence for FLE. ... Methinketh there is a flaw in the FLE (or three) here ... First, what is your evidence for "deep homology" and how is this specifically predicted by FLE as opposed to being a post hoc ergo propter hoc observation? (ie -- if it is something known before your hypothesis was developed, then it would be, should be, relegated to the objective evidence used to form the hypothesis not to test it). You need to predict something unknown and then test for it. Second, how do you know that "deep homology" is not predicted by FLU or NFLU? (ie-- by "natural (FLU/NFLU) laws"). Just stating that it isn't does not meet scientific criteria.
The process of Microevolution involves changes in the composition of hereditary traits, and changes to the frequency of their distributions within breeding populations from generation to generation, in response to ecological challenges and opportunities. The process of Divergent Speciation involves the division of a parent population into two or more reproductively isolated daughter populations, which then are free to (micro) evolve independently of each other. The Theory of Evolution (ToE), stated in simple terms, is that the process of Microevolution, and the process of Divergent Speciation, are sufficient to explain the diversity of life as we know it, from the fossil record, from the genetic record, from the historic record, and from everyday record of the life we observe in the world all around us. The ToE predicts that divergent speciation results in a pattern of nested hierarchies of descent from common ancestor breeding populations, of clades within clades, and this pattern necessarily results in "deep homologies" within the branches of each of the clades, and the deeper you go into the nested hierarchy ancestry the deeper you go into the "deep homologies" that are present.
| ^ a / \ / \ / \ / ^ b / / \ / / \ e d c Homologies shared by a, b, c, d,and e are "deep homologies" predicted by ToE. Thus the ToE predicts "deep homologies" necessarily develop from microevolution plus divergent speciation. The ToE also predicts that "deep homologies" shared by a and e but not by b will also not be shared by c and d. This pattern is not predicted by FLE, as I understand it, but is observed to occur. Can you explain this apparent omission? Third, the ToE also predicts derived traits, where a trait from an ancestral population is modified via microevolution into a variation that does not occur in the parent population. The ToE predicts that these derived traits will be homologous in descendant populations. Thus b will have derived traits that are not shared with a (or e) but which are shared with c and d AND that these derived traits have predecessors in a that have been modified in b, rather than appearing de novo. Does your FLE predict this? Then there is the issue of convergent evolution: why take two paths to reach a single front loaded result? Is this predicted?
I'm afraid that that's not how you go about gathering evidence for a scientific hypothesis. ... Curiously, I've only been told this by people pushing their beliefs as an hypothesis, rather than people talking about scientific hypothesis and how they are properly tested. Amusingly, they also tried to use evidence used to develop an hypothesis as evidence that it was correct.
... If scientific hypothesis A predicts x-y-z, while hypothesis B does not predict x-y-z but could explain the occurrence of x-y-z, the occurrence of x-y-z is evidence for hypothesis A. ... No, it is just evidence that the hypothesis is not invalidated, a seemingly minor but important distinction, and that it has not been tested to differentiate it from hypothesis B. Particularly if you have not show that it is not predicted by hypothesis B. Similarly if hypothesis A predicts x-y-q does NOT occur, and B does not predict the occurrence of x-y-q then the non-existence of x-y-q neither confirms nor invalidates hypothesis A or B.
... Case in point: the competing hypotheses concerning the origin of the type III secretion system (TTSS). The "sister group" hypothesis suggests that the TTSS and the flagellum are monophyletic, sharing a common ancestor, while the "flagellum first" model posits that the TTSS arose directly from the flagellar system. Confirmation of a prediction of either of these hypotheses would be evidence for that hypothesis, regardless of whether the other hypothesis could explain the observation behind the confirmed prediction. ... No, it would just confirm that either hypothesis could be correct. You need a prediction that occurs by A and not by B or one that occurs by B and not by A in order to differentiate which hypothesis is a better explanation of the evidence. The purpose of an hypothesis (and a theory) is to explain the evidence, so as long as B explains the evidence it is a valid (not invalidate) theory. Consider where each hypothesis predicts something the other does not, but does not rule out. Both predictions come true: which one is better than the other in explaining the evidence?
... So, IMHO, your above question assumes we must be able to rule out non-telic mechanisms, when we do not. ... In your opinion. Curiously, opinion has been shown to have little effect on reality. To be a proper test of an hypothesis you need to predict something that cannot occur by the other hypothesis or vice versa. If we assume A and derive B from A, and B predicts C occurs, then occurrence of C is evidence for B If we assume notA and derive D from notA, and D does not prevent C from occurring, then the occurrence of C is not evidence against D Does A or notA exist? If you cannot tell, then the occurrence of C is not evidence that B is any more likely to be true than not true. Enjoy. Edited by RAZD, : added derived traitsby our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||
|
Genomicus Member (Idle past 1960 days)  Posts: 852 Joined: |
What we don't know is how that life got there. Your delivery system requires a previously developed life\being while mine only requires observed systems for delivery of observed materials from space to the surface of a planet. With no evidence of your delivery system you must admit that this is an extremely weak link in your hypothesis: you have to assume that it existed. But this part of your thesis is not that different from plain ole' panspermia, where meteorites or other cosmic bodies deliver already-formed life on our planet. The delivery system of non-directed panspermia relies on observed systems for delivery of life. Directed panspermia involves an intelligence. The phrase "extremely weak link" is a subjective one, is it not?
You forget the age of these interstellar clouds - because of their distance from earth what we are seeing were there before the earth formed. Thus the absence of identifiable interstellar cells is problematic for a panspermia concept. See here: F. Hoyle and C. Wickramasinghe. On the Nature of Interstellar Grains, Astrophysics and Space Science, 1979. They found evidence of freeze-dried bacteria and algae in outer space.
So you would agree then that there is nothing special about life existing on earth? Special is a subjective term. To me, life is extraordinarily special regardless of its origin.
And yet you are still only copying an existing system for making life. The FLE hypothesis does not address the origin of life. Your thesis does. That's a fundamental difference between our two viewpoints.
Being able to construct life from pre-biotic materials would be a prediction of FLU but not of FLE, yes? Does FLE predict that this will not be possible? It certainly seems to imply it to me, otherwise why is FLE necessary? Why on earth would FLE predict this to not to be possible? If one was able to construct life from pre-biotic materials, it would have little relevance to the validity (or lack thereof) of the FLE hypothesis. It wouldn't matter either way for the FLE hypothesis, since the FLE hypothesis is not a hypothesis about the origin of life. It is a hypothesis about the development of life here on earth, regardless of life's ultimate origin.
A scientific hypothesis, as opposed to a philosophical one, is based on objective evidence or objective observation (rather than on personal intuition: see Percy Message 210), AND it is testable. The FLE hypothesis is based on objective evidence and it is testable. We find evidence of key eukaryotic proteins sharing deep homology with functional but unnecessary prokaryotic proteins. This is not expected from conventional evolutionary theory, as I explained here:
I argue that the FLH predicts that proteins of major importance in eukaryotes and advanced multi-cellular life forms (e.g., animals, plants) will share deep homology with proteins in prokaryotes. I have discussed this prediction with various critics of the FLH, and the most common objection seems to be that non-teleological evolution also makes this prediction. I disagree, so let me explain. Life seems to require a minimum of about 250 genes (Koonin, Eugene V. How Many Genes Can Make a Cell: The Minimal-Gene-Set Concept, 2002. Annual Reviews Collection, NCBI) — a proto-cell would not require that many genes. Thus, it would be perfectly acceptable, under the non-teleological model, that the last common ancestor of all life forms had approximately 250 genes, add or take a few. From this small genome, gene duplication events would have occurred, subsequently followed with mutations in the new genes, would lead to a novel protein. Over time, then, and through gene and genome duplication/random mutation, this small genome would evolve into larger genomes. This model is perfectly acceptable with the non-teleological hypothesis, and the non-teleological hypothesis does not predict otherwise. However, this model — where a minimum genome gradually evolves into the biological complexity we see today, through gene duplication, genome duplication, natural selection, and random mutation — is not compatible with the front-loading hypothesis. This is because front-loading requires that the first genomes have genes that would be used by later, more complex life forms. Of the 250 or so genes required by life, none of them could encode proteins that would be used later in multicellular life forms (excluding the proteins that are necessary to all life forms). A front-loading designer couldn’t possibly hope to stack the deck in favor of the appearance of plants and animals, for example, by starting out with a minimal genome. Look at it this way. With a minimal genome of 250 genes that are involved in metabolism, transcription, translation, replication, etc., evolution could tinker with that genome in any way imaginable, so that you couldn’t really front-load anything at all with a minimal genome. You couldn’t anticipate the rise of animals and plants. Such a genome would not shape subsequent evolution. If the last common ancestor of all life forms had a minimal genome, and if you ran the tape of life back, and then played it again, a totally different course of evolution would result. But if you loaded LUCA with genes that could be used by animals and plants, you could predict that something analogous to animals and plants would arise. If you loaded this genome with hemoglobin, rhodopsin, tubulin, actin, epidermal growth factors, etc. — or homologs of these proteins — something analogous to animal life forms would probably result over deep-time. Given that you couldn’t really front-load anything with a minimal genome consisting of about 250 genes, under the front-loading hypothesis, it is necessary that LUCA contain unnecessary (but beneficial) genes that would later be exploited by more complex life forms. Non-teleological evolution does not require this. It has no goal, unlike front-loading. It tinkers with what is there — and if a minimal genome was all that was there, it would tinker around, eventually producing endless forms most beautiful as Darwin so famously put it. On the other hand, front-loading is goal-oriented: a minimal genome does not allow one to plan the origin of specific biological objectives. Thus, under the front-loading hypothesis, we would predict that important proteins in eukaryotes, animals, and plants will share deep homology with unnecessary but functional proteins in prokaryotes. Non-teleological evolution does not predict this. Non-teleological evolution could explain that observation, but it does not predict this. And this is the important point to understand. There is nothing in non-teleological evolution that requires multi-cellular proteins to share deep homology with unnecessary prokaryotic proteins — but front-loading demands this. There is nothing in non-teleological evolution that requires that LUCA have a genome larger than the minimum genome size — but for front-loading to occur, this must be the case. I conclude, then, that this prediction is made by the front-loading hypothesis, but it is not made by non-teleological evolution, and so front-loading is certainly testable. Curiously, I'm pointing out that your hypothesis is not testable - or at least you have not demonstrated how it could be properly tested. I certainly have. Here:
It is important for a hypothesis to make testable predictions. Here, I will try to briefly describe how the FLE hypothesis makes predictions that are not made by conventional theory. Before beginning, however, I would like to point out that, in this thread, I do not intend to discuss in depth the issue of whether some of these predictions have, in fact, been confirmed. In this thread, I am primarily interested in discussing if these predictions differ from those generated by conventional theory. Let me begin with a prediction concerning the origin of molecular machines like cilia. Intra-flagellar transport (IFT) particles are involved in ciliary function in most eukaryotes. These proteins contribute to ciliary function, and any eukaryotes that lack these IFT proteins — such as Plasmodium -- are probably degenerate cilia and do not represent the structure of the last cilia common ancestor. The point is this: under the non-teleological framework, co-option events are primarily responsible for the origin of this motility organelle and its IFT proteins. Under this model, random co-option events of proteins in the cell resulted in the functional association of different proteins, which would have been preserved by natural selection — and over time, through repeating this step, finally a cilium arose. This is, in essence, the non-teleological hypothesis for the origin of the eukaryotic flagellum. Given that the existence of Metazoa seems to require the existence of cilia, under the FLE model, cilia were front-loaded. How would cilia be front-loaded? The FLE hypothesis is only at its beginning stages, so one should not expect, at the present moment, a rigorous FLE model for the origin of the cilium. However, I can offer a cursory model for the FLE origin of the cilium. In this model, the first genomes would be designed with components that would later be used by the cilium. In other words, homologs of the core, necessary IFT proteins would be designed into the first genomes. They’d be given a function, such that their basic 3D shape is conserved over deep-time. If they were given a function where their 3D shape would be substantially changed over deep-time, then the front-loading designer couldn’t possibly hope that when these proteins associated, their shapes would complement each other correctly such that a cilium could arise. From here, we can develop our FLE prediction. The non-telic hypothesis for the origin of the cilium does not require or predict that the prokaryotic homologs of IFT proteins be well-conserved in sequence identity. In fact, it’s certainly possible that the non-telic hypothesis predicts that most of the prokaryotic homologs of the core IFT proteins will be loosely conserved in sequence identity: a protein that is not under stringent functional constraints will be more likely to be co-opted into a novel role by chance without being deleterious. For example, H4 histone is one of the most highly conserved proteins in eukaryotes. To me, at least, it seems that it would be much more likely that if H4 histone was duplicated and then co-opted into an entirely novel function a non-adaptive effect would occur than if a fibrinopeptide, for example (which are not at all highly conserved), were co-opted into this novel role. This would be an interesting line of research, but I don’t intend to explore this argument further, because the fact remains: the non-telic hypothesis for the origin of the cilium does not require or predict that the prokaryotic homologs of core IFT proteins be well-conserved in sequence identity, while the FLE hypothesis for the origin of the cilium predicts that the prokaryotic homologs of core IFT proteins would be well-conserved in sequence identity, more so than the average prokaryotic protein. This is a testable prediction: we would need to find a prokaryotic homolog of a core IFT protein, and then conduct pairwise comparisons of that IFT homolog with its prokaryotic orthologs, and check its degree of sequence conservation. There is nothing in the non-telic hypothesis that predicts this hypothetical prokaryotic homolog will be highly conserved in sequence identity, more so than the average prokaryotic protein. You will not find anything like this prediction in the scientific literature. But you have to admit that it is not a scientific hypothesis, but an assumption based on personal intuition\opinion\wish\hope, and that your FLE hypothesis is founded on that assumption. But there is evidence for front-loaded evolution. See above.
If the prediction is for something that cannot differentiate FLE from FLU or NFLU, then confirmation of a prediction just means that it is not falsified, while the validity of the hypothesis is not tested. I disagree, based on several considerations (such as the discussion of the origin of the TTSS in the scientific community). For example, evolutionary theory predicts a nested hierarchical pattern. Can creationism explain this observation? You bet it can. It merely needs to say that goddunnit. Thus, based on your logic, evolutionary theory has not been tested by the existence of an NHP; it simply has not been falsified.
First, what is your evidence for "deep homology" and how is this specifically predicted by FLE as opposed to being a post hoc ergo propter hoc observation? (ie -- if it is something known before your hypothesis was developed, then it would be, should be, relegated to the objective evidence used to form the hypothesis not to test it). You need to predict something unknown and then test for it. I have explained above how it is specifically predicted by FLE. It has been confirmed for a number of proteins (objective evidence for FLE), but it has yet to be confirmed for other proteins (a test for the FLE model). Then there is the example from IFT particles in cilia, which is a test for the FLE hypothesis.
Second, how do you know that "deep homology" is not predicted by FLU or NFLU? (ie-- by "natural (FLU/NFLU) laws"). Just stating that it isn't does not meet scientific criteria. See above where I explain why FLE predicts "deep homology" with unnecessary but functional proteins in prokaryotes, while other conventional theory do not. There is nothing in NFLU that would predict this, either.
The ToE predicts that divergent speciation results in a pattern of nested hierarchies of descent from common ancestor breeding populations, of clades within clades, and this pattern necessarily results in "deep homologies" within the branches of each of the clades, and the deeper you go into the nested hierarchy ancestry the deeper you go into the "deep homologies" that are present. Again, see above where I explain why ToE does not predict the specific pattern of deep homology that I am referring to.
This pattern is not predicted by FLE... Ummm, why do you think that?
No, it is just evidence that the hypothesis is not invalidated, a seemingly minor but important distinction, and that it has not been tested to differentiate it from hypothesis B. Particularly if you have not show that it is not predicted by hypothesis B. But if hypothesis B does not make that prediction, while A does, then confirmation of that prediction is evidence for A, even if B can explain it through ad hoc rationalizations.
To be a proper test of an hypothesis you need to predict something that cannot occur by the other hypothesis or vice versa. Really? The "flagellum first" hypothesis for the origin of the TTSS predicts that the flagellum will be more widely distributed among bacteria phyla than the TTSS. The "sister group" hypothesis could explain this observation (through lateral gene transfer) but it does not predict it. Thus, confirmation of the above prediction is evidence in favor of the "flagellum first" hypothesis. That's how investigators of the origin of the TTSS see it, and that's how I see it. Also: if we go by your argument, then the NHP is not evidence for common descent, since the appearance of the NHP can also occur under the creationism model (where goddunnit), even though the creationism model does not predict it. Edited by Genomicus, : No reason given.
|
|||||||||||||||||||||||||
|
RAZD Member (Idle past 1424 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Genomicus, excuse the length - we may need to break this down into subtopics if this goes on growing.
Please note that I added a section on derived traits and a question about convergent evolution before I saw your reply:
Message 211: Third, the ToE also predicts derived traits, where a trait from an ancestral population is modified via microevolution into a variation that does not occur in the parent population. The ToE predicts that these derived traits will be homologous in descendant populations. Thus b will have derived traits that are not shared with a (or e) but which are shared with c and d AND that these derived traits have predecessors in a that have been modified in b, rather than appearing de novo. Does your FLE predict this? Then there is the issue of convergent evolution: why take two paths to reach a single front loaded result? Is this predicted? But this part of your thesis is not that different from plain ole' panspermia, where meteorites or other cosmic bodies deliver already-formed life on our planet. The delivery system of non-directed panspermia relies on observed systems for delivery of life. Directed panspermia involves an intelligence. The phrase "extremely weak link" is a subjective one, is it not? The weak link is your completely unevidenced assumption of an intelligence and that the process is directed and designed to front-load the system, and in this your thesis is considerably different from "plain ole' panspermia" or FLU ... PLUS your FLE relies entirely on this being true: that is your extremely weak link.
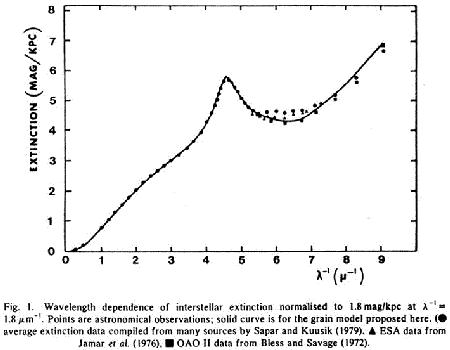
See here: F. Hoyle and C. Wickramasinghe. On the Nature of Interstellar Grains, Astrophysics and Space Science, 1979. They found evidence of freeze-dried bacteria and algae in outer space. Been there, hence my comment being what it was.
quote: They are not positively identified as interstellar cells or the remains of them. The possibility of constructs like buckyballs were not known by Hoyle and Wickramasinghe when they published in the 70's:
quote: And likely many other patterns are possible for forming hollow shells, with the ones observed being more stable. I would expect that a dodecahedron pattern would also be stable and larger geodesic shapes may also be possible. The graph used by Hoyle and Wickramasinghe
As NASA's recent evidence for buckyballs is the same kind of evidence as the 40+ year old evidence used by Hoyle and Wickramasinghe, and is much more discrete in analyzing the signal (two peaks instead of one), my money is on fullerenes rather than cells as the more accurate explanation. Furthermore, fullerenes are now known to clump together into larger particles:
quote: Stacked like a crystalline structure using buckyballs as the matrix molecules ... could we see superbuckyballs made of buckyballs? In addition, there are other problems for them being dried shells of bacteria: they are not necessarily living organisms (or spores) nor do they necessarily have functional DNA inside - the pattern is for hollow shells, not living cells --- that has to be assumed for your thesis. Do a google on buckyball panspermia and see how many advocates are trying to cram panspermia inside the buckyballs.
The FLE hypothesis does not address the origin of life. Your thesis does. That's a fundamental difference between our two viewpoints. Yes, it makes mine more complete and comprehensive, while yours still rests on certain incompletely defined assumptions being true without any objective evidence in this area.
Why on earth would FLE predict this to not to be possible? If one was able to construct life from pre-biotic materials, it would have little relevance to the validity (or lack thereof) of the FLE hypothesis. ... Because (a) otherwise the FLE is unnecessary and (b) the inability to produce results would tend to invalidate the concept of abiogenesis by "natural (FLU/NFLU) laws" - leaving the field open for life being "salted" on earth.
... the FLE hypothesis is not a hypothesis about the origin of life. It is a hypothesis about the development of life here on earth, regardless of life's ultimate origin. But it makes assumptions about the origin of the cells and their delivery to the earth, assumptions that are just assumed to be true without testing or validation.
The FLE hypothesis is based on objective evidence and it is testable. We find evidence of key eukaryotic proteins sharing deep homology with functional but unnecessary prokaryotic proteins. This is not expected from conventional evolutionary theory, as I explained here:
{this is either poorly explained, I may have failed to understand it properly, or it is just post hoc ergo propter hoc circular reasoning. It certainly seems to use a lot of verbiage for what is claimed to be a simple concept ... let me know if you can simplify it. It also seems to rely heavily on some basic assumptions (genome size for example) and conflation of LUCA with the smallest possible life form. Also see The Surprising Complexity of LUCA. There is nothing in the definition of the Last Universal Common Ancestor (LUCA) that says it must be the simplest or the smallest form of life.} First, we may see evidence in life occurring today of "key eukaryotic proteins sharing deep homology with functional but unnecessary prokaryotic proteins." Seeing as all these forms are current living organisms they do not necessarily make any claim about early life. We also know that unicellular life is rife with horizontal transfer of material from one organism to another, even across the designations of the major domains of life (the cells don't care what they are). The laws of an FLU would allow this horizontal transfer, but it should be completely unnecessary and certainly is not predicted by an FLE that has a complete package in the initial life.
quote: Curiously, I would expect that the FLE would predict a single unequivocal tree structure at the roots of life without Horizontal Gene Transfer (HGT), including genes for the production of proteins. Second, proteins are proteins, they don't depend on any specific branch of life to exist, they just need to be combined from amino acids in roughly the same order by any type of cell so we would expect (deep) homologies from shared ancestors along with some variation accumulated over time. If they serve little or no function in the fully developed cell at this stage in the game of existence (3.5 billion years after the first life) we can assume that some variation would occur, such that the proteins are not as homologous across domains as they are within domains.
quote: Similar but different, just as should be expected. Note also that we have {p, b, a}, {p, b, nota}, {p, notb, a}, and {notp, b, a} homologies, which looks a lot more like horizontal transfer than front-loading to me.
I certainly have. Here:
{again this is either poorly explained, I may have failed to understand it properly, or it is just post hoc ergo propter hoc circular reasoning. It certainly seems to use a lot of verbiage for what is claimed to be a simple concept ... let me know if you can simplify it. Your predictions seem to be that things we observe are due to FLE rather than NFLE ("non-telic") processes. In order for this to be a prediction for FLE you need to show that they could not occur under NFLE, and you haven't: this is NOT a test, as it does not tell us that FLE occurred instead of NFLE.} Curiously, you appear to using evidence that is known prior to the hypothesis formation as a validation test, when this is properly included in your review of the current state of knowledge in forming an hypothesis in order to explain all the known objective empirical evidence. That is not a true prediction of the hypothesis, instead it is a demonstration of consilience between the known objective evidence and the hypothesis. A proper prediction is for something not already known, something new, and preferably something that cannot occur without the hypothesis or theory.
But there is evidence for front-loaded evolution. See above. But there isn't -- see above. There is objective evidence that cilia exist. The proper approach is to take your example of your purported FLE instance (cilia exist), show that this could not occur in a NFLE world, and then form your your hypothesis test. Unfortunately, for you, you have done the opposite - shown how the NFLE would work.
I disagree, based on several considerations (such as the discussion of the origin of the TTSS in the scientific community). For example, evolutionary theory predicts a nested hierarchical pattern. Can creationism explain this observation? You bet it can. It merely needs to say that goddunnit. Thus, based on your logic, evolutionary theory has not been tested by the existence of an NHP; it simply has not been falsified. Disagree all you want, but show me the evidence. You're going to have to provide me with more information than this hopefully from professional journals such as you have already used for this "discussion" you have asserted. I'm betting it is about details rather than yes or no.
I have explained above how it is specifically predicted by FLE. It has been confirmed for a number of proteins (objective evidence for FLE), but it has yet to be confirmed for other proteins (a test for the FLE model). Then there is the example from IFT particles in cilia, which is a test for the FLE hypothesis. Except that these are, as already noted, predicted by ToE and our current knowledge of early life and horizontal transfer among unicellular life. In no case have you provided evidence of something predicted that was not previously known. That is not a prediction, it is a claim of consilience between the known objective evidence and the hypothesis. Seeing as any new hypothesis must explain all current known existing evidence, demonstration of consilience with that evidence is not the result of prediction, rather this is a necessary part of the effort, the homework, that needs to go into the formation of a new hypothesis - checking to see that it is consilient with all known data. What you need to predict is something not already known, something new, and preferably something that cannot occur without FLE.
See above where I explain why FLE predicts "deep homology" with unnecessary but functional proteins in prokaryotes, while other conventional theory do not. There is nothing in NFLU that would predict this, either. See above where I explain how "conventional theory" does in fact predict "deep homologies" ... (and derived traits in the added sections of the previous message noted above, along with the question of convergent evolution). Curiously, the ToE makes no prediction that preserved elements need to be functional or necessary, just that they not be deadly.
Again, see above where I explain why ToE does not predict the specific pattern of deep homology that I am referring to. Again, see above where I explain why ToE does predict the patterns of deep homologies regardless of how specific they are. You have not shown a specific instance of any element that cannot occur by "conventional theory" and in fact have said so ("This is, in essence, the non-teleological hypothesis for the origin of the eukaryotic flagellum").
Ummm, why do you think that? If you can then show me how. I'm open to evidence based logical argument.
But if hypothesis B does not make that prediction, while A does, then confirmation of that prediction is evidence for A, even if B can explain it through ad hoc rationalizations. Repeating your assertion does not make it any more valid. Using A you make a prediction that you have not made by BUsing B you make a prediction that you have not made by A Both predictions are true So which is more valid? The function of a hypothesis or a theory is to explain the evidence, it doesn't matter if the evidence is new or old, predicted or covered by the mechanics that went into the development of the hypothesis or theory. The function of a prediction test is to predict something not already known, something new, and preferably something that cannot occur without the hypothesis or theory.
Really? The "flagellum first" hypothesis for the origin of the TTSS predicts that the flagellum will be more widely distributed among bacteria phyla than the TTSS. The "sister group" hypothesis could explain this observation (through lateral gene transfer) but it does not predict it. Thus, confirmation of the above prediction is evidence in favor of the "flagellum first" hypothesis. That's how investigators of the origin of the TTSS see it, and that's how I see it. Really? A "flagellum second" hypothesis would be a counter hypothesis (antithesis) to the "flagellum first" hypothesis and it would predict that TTSS would be more prevalent than the flagellum, which would be a proper falsification test for the "flagellum first" hypothesis. This test failed to falsify "flagellum first" -- it's simple cladistics:
| ^ a = flagellum / \ / \ / ^ b = TTSS / / \ / / \ This is why I talk about having a notA prediction to validate your hypothesis. Passing this test (and others as the testing progresses) makes the hypothesis stronger than not being tested. It doesn't make it true, as no amount of testing can accomplish that, and it doesn't make other hypothesis invalid unless they are mutually exclusive. What horizontal gene transfer would predict is that both the flagellum and TTSS could be transfered via HGT from bacteria that posses these trait to ones that don't. Natural selection says that these with the transfered gene would be preserved if they expressed the gene and it provided survival and reproductive benefit. This would predict that we would have some "flagellum first" and some "flagellum second" bacteria (and that this could muddy the evidence). It would also predict that all the species of bacteria where the gene originated would have the gene but not all species of bacteria that received the transfer would (however, those 'with' would likely be classified as a new species that would all have the gene). Thus these hypothesis are not incompatible or exclusive, and both hypothesis could be working at the same time. HGT is not invalidated here as the explanation (and it has been validated elsewhere as occurring), and it's prediction is not tested. The true answer probably lies with a combination of both hypothesis to explain all the evidence. This puts the "sister group" hypothesis in the same position as String Theory (really an hypothesis) compared to the "standard model" in physics, ... and your hypothesis. An hypothesis that is tested by passing invalidation tests is stronger than one that has not. This applies to "the standard model" the "flagellum first" hypothesis, common descent and the ToE, but it does not apply to String Theory, the "sister group" hypothesis, ... or your hypothesis. Your hypothesis is still not tested by a proper prediction. You can't predict something already known to exist, you can just show how the hypothesis explains it.
Also: if we go by your argument, then the NHP(1) is not evidence for common descent, ... Curiously, that's not what I said. What I said was that
To be a proper test of an hypothesis you need to predict something that cannot occur by the other hypothesis or vice versa. A falsification test for common ancestry is the genetic data: specifically there should be no ERV's of the same kind in the same locations in the genes. The genetic analysis not only shows these occurring, but occurring with "deep homologies" in a nested clade hierarchy pattern, just as predicted by the ToE. You don't have a proper invalidation test, and you don't have a prediction for something not already known, something new, and preferably something that cannot occur without the hypothesis or theory.
... since the appearance of the NHP(1) can also occur under the creationism model (where goddunnit), even though the creationism model does not predict it. Except that (a) the "creationism model" pretends to do science after "special creation" rather than whole sale tweaking of every piece of evidence (a position they are forced into by the increasing evidence for ToE), (b) that the existence of NHP alone does not explain the existence of ERV sharing, and when the "goddunnit" card gets played every time the "creationism model" runs into problems, then it is constantly being revised to include more "goddunnit" and less natural processes validating the model. This means overall validation by objective evidence is shrinking rather than growing, and (c) invalidation of one hypothesis or theory does not make a single other hypothesis or theory more valid. Enjoy (1) - NHP as in Non-Human Primate?by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||
|
Genomicus Member (Idle past 1960 days) Posts: 852 Joined: |
With respect RAZD, I have a feeling that you might possibly be misunderstanding the FLE hypothesis, and you seem to be under the impression that the FLU is completely incompatible with FLE.
And you're making kinda suspicious statements sometimes. For example, I said:
The FLE hypothesis does not address the origin of life. Your thesis does. That's a fundamental difference between our two viewpoints. To which you replied:
Yes, it makes mine more complete and comprehensive, while yours still rests on certain incompletely defined assumptions being true without any objective evidence in this area. But my focus does not lie in the origin of life as much as in the development of life on earth. And I think you need to understand that. I know you're very interested in the origin of life and the origin of natural laws, but I'm not. So when you say it makes your thesis more complete, my answer is: so what? I am interested in the development of life on earth, and not the things you are interested in. How life originated has little relevance to the FLE hypothesis (unless one is going to argue that life evolved here, on our planet). Here's another interesting statement you made:I said: Why on earth would FLE predict this to not to be possible? If one was able to construct life from pre-biotic materials, it would have little relevance to the validity (or lack thereof) of the FLE hypothesis. ... And you said:
Because (a) otherwise the FLE is unnecessary and (b) the inability to produce results would tend to invalidate the concept of abiogenesis by "natural (FLU/NFLU) laws" - leaving the field open for life being "salted" on earth. But you fail to realize that the FLE hypothesis does not address the origin of life. You're trying to force it to do so, hence your claim that the construction of life from pre-biotic chemicals would go against the FLE hypothesis. But it wouldn't, since how life originated doesn't matter to the FLE hypothesis. There's a lot of stuff in your comment that I really haven't got the time to respond to. I will, however, clarify the "deep homology" prediction of the FLE hypothesis, since for some reason it isn't clear to you: The non-telic hypothesis for the evolution of life on earth makes no predictions regarding the genome size of the LUCA. But the FLE hypothesis requires that the LUCA have unnecessary but functional genes that would later be used by complex life forms. Thus, the FLE hypothesis requires that the LUCA be unnecessarily complex. Non-teleological evolution does not have this requirement; nor does it make any predictions regarding the complexity of the LUCA. It also seems to me that you completely misunderstood the prediction regarding IFT particles and the cilium.
There is objective evidence that cilia exist. The proper approach is to take your example of your purported FLE instance (cilia exist), show that this could not occur in a NFLE world... But that wasn't the prediction of the FLE hypothesis. Of course cilia exist, and its existence isn't the issue here. It's the level of sequence conservation among prokaryotic homologs of ciliary components. Next, you make the very, very, very strange claim that horizontal gene transfer is somehow not expected by FLE - even though horizontal gene transfer is an extremely good mechanism for forwarding designs into the future. You make the following claim:
Seeing as all these forms are current living organisms they do not necessarily make any claim about early life. We also know that unicellular life is rife with horizontal transfer of material from one organism to another, even across the designations of the major domains of life (the cells don't care what they are). But we can test for horizontal gene transfer, and this is a fatal flaw in your above argument (you don't even mention this issue). We can construct a phylogenetic tree of the gene in question, and compare it to 16s rRNA phylogenies to see if they are congruent. If they are not, then HGT is likely at play here. But if they are, then HGT is almost certainly not primarily responsible for the resulting tree. Yes, I know this isn't at all a reply to many of your points. But before even thinking about replying to your other points, I want to make something clear here. You seem to be under the idea that somehow the FLU and the FLE are at odds, and incompatible. But why do you think that? And, frankly, I don't really know how this thread - which was originally supposed to be about front-loaded evolution - got sidetracked into a discussion on your thesis of FLU. I have no problem with you making issues of the testability etc. of the FLE, but I don't really see any reason to discuss your FLU ideas in this thread. Half of our responses to each other seem to be about the FLU, when I wouldn't care either way. Hopefully, I'm not coming across as snarky or rude, 'cause I don't mean to be. I'm just saying... Edited by Genomicus, : No reason given.
|
|||||||||||||||||||||||||
|
RAZD Member (Idle past 1424 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Genomicus,
With respect RAZD, I have a feeling that you might possibly be misunderstanding the FLE hypothesis, ... Always a possibility. Let me see if I can state it properly and you can correct me where I am wrong:
premise 1: aliens of vast intelligence (or god/s) designed a cell premise 2: the cell was designed to be front-loaded with genes that would bias (direct) any evolution of life from that cell premise 3: aliens of vast intelligence (or god/s) put copies of this cell in space premise 4: one or more of these cells were delivered to planetary surfaces, including earth, via meteors and the like, where they then started reproducing and evolving hypothesis: the bias (direction) built in to the first cells has guided all subsequent evolution of life on earth; evolution on earth has been biased (guided) to evolve along the pathways necessary to arrive at the diversity of life we currently see on earth by these front-loaded cells. Is that a fair assessment?
But my focus does not lie in the origin of life as much as in the development of life on earth. ... And yet your hypothesis is unable to remove a single premise above without falling to pieces. It does not matter whether these premises are stated or unstated, as each one is critical to the formation of the front-loading element in your hypothesis. Remove front-loading from your hypothesis and it reverts to pansperia as an explanation for first life, from which all life subsequently evolved by natural (FLU or NFLU) laws. You need all those premises included before you can get the cells to be front-loaded in your hypothesis. The premises aren't there to explain panspermia, OOL, or evolution, but to explain the front-loading of your hypothesis.
And I think you need to understand that. I know you're very interested in the origin of life and the origin of natural laws, but I'm not. So when you say it makes your thesis more complete, my answer is: so what? I am interested in the development of life on earth, and not the things you are interested in. How life originated has little relevance to the FLE hypothesis (unless one is going to argue that life evolved here, on our planet). But how the front-loading originated is of critical importance to your FLE hypothesis. Let us posit that the appearance of front-loaded cell life was through the action of purely naturalistic (whether FLU or NFLU) laws, then evolution would be naturally' biased rather than design biased, yes? We can call this the NFLE hypothesis (antithesis). Thus to posit a design element, you need to posit a source for that design, and a method to realize the design and you need a means to test for it against the NFLE.
But you fail to realize that the FLE hypothesis does not address the origin of life. You're trying to force it to do so, hence your claim that the construction of life from pre-biotic chemicals would go against the FLE hypothesis. But it wouldn't, since how life originated doesn't matter to the FLE hypothesis. The question is how the front-loading originated. If we assume the NFLE hypothesis, then this would require life originating from some other source, one of which could be the construction of life from pre-biotic chemicals, and evidence of this being possible would be evidence for the NFLE rather than the FLE. I'm following your hypothesis to logical conclusions: if FLE is responsible for life on earth then no other mechanism for origins should exist.
There's a lot of stuff in your comment that I really haven't got the time to respond to. I will, however, clarify the "deep homology" prediction of the FLE hypothesis, since for some reason it isn't clear to you: The non-telic hypothesis for the evolution of life on earth makes no predictions regarding the genome size of the LUCA. But the FLE hypothesis requires that the LUCA have unnecessary but functional genes that would later be used by complex life forms. Thus, the FLE hypothesis requires that the LUCA be unnecessarily complex. Non-teleological evolution does not have this requirement; nor does it make any predictions regarding the complexity of the LUCA. The NFLE makes no prediction regarding the genome size or complexity of LUCA, it could be large or small. Confirmation that it is large and complex does not rule out NFLE, thus such a prediction does not differentiate FLE from NFLE. If if provides anything it provides for confirmation bias rather than open-minded skeptical validation. Furthermore, a smaller size and complexity of LUCA does not rule out the existence of "deep homologies" existing in the offspring of LUCA.
... And, frankly, I don't really know how this thread - which was originally supposed to be about front-loaded evolution - got sidetracked into a discussion on your thesis of FLU. I have no problem with you making issues of the testability etc. of the FLE, but I don't really see any reason to discuss your FLU ideas in this thread. Half of our responses to each other seem to be about the FLU, when I wouldn't care either way. Whether evolution is via FLE or NFLE is not important to the FLU, and the special FL design of the FL cell is not necessary to the FLU. If you don't want to discuss FLU aspects, the we can concentrate on the differences between your FLE and a NFLE hypothesis, and how your testing proposes to differentiate one from the other. In NFLE we don't have a front-loaded cell, so evolution occurs purely by NFLE means and processes, including evolution as currently defined and used in science:
(1) The process of Evolution involves the change in the frequency distribution and composition of hereditary traits within breeding populations from generation to generation, in response to ecological challenges and opportunities. This means that from generation to generation many traits are preserved within the populations (even if their frequency distribution changes), but that some may be altered and adapted. This means that there would be homologous traits preserved from generation to generation, and there would also be the appearance of derived traits (that can then become homologous in following generations). Thus the NFLE does not rule out deep homologies which you claim FLE predicts, and it also predicts deep derived traits that can then become deep homologies in later generations. As far as I can see your FLE does not predict deep derived traits nor do you have a test to separate original deep homologies from ones from deep derived traits and I also have to ask if deep derived traits are not counter to your front-loading hypothesis (as they should not be necessary if everything necessary was provided in the original front-loaded cells, yes?). Thus deep derived traits would seem to be a falsification test for FLE, yes?
| | a / \ / \ / g b / \ / \ / \ / | | h c | | / \ / \ | | / i / \ | | | / \ d e f j k l m Shared homologies in all populations (a thru m) would be deep homologies occurring via NFLE evolution. Shared homologies only in populations b thru f would also be deep homologies occurring via NFLE evolution from the deep derived traits in b that separate b from g, while shared homologies only in populations g thru m would also be deep homologies occurring via NFLE evolution from the deep derived traits in g that separate g from b. Curiously, it is just this pattern of deep deep homologies and deep derived traits that form the nested clade hierarchy pattern predicted by evolution in a NFLE world. For an FLE hypothesis I would expect to see homologies between b and g for unused copies of what otherwise appear to be deep derived traits, and logically this would also apply for all other appearances of derived traits in all later populations (ie no trait is ever derived, just expressed and used or not expressed and not used). This not only means a large and complex LUCA, as you state, but a large and complex descendant at every stage of evolution.
Hopefully, I'm not coming across as snarky or rude, 'cause I don't mean to be. I'm just saying... Likewise. I'm just trying to explore what differentiates your FLE from a NFLE, in order to better understand your hypothesis, the tests you have proposed, and what the evidence actually shows. Let me know where I've gone wrong eh? Enjoy Edited by RAZD, : ngls, subtitle Edited by RAZD, : clrty Edited by RAZD, : trying to remove curiously occurring extra spacingby our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||
|
Taq Member  Posts: 10028 Joined: Member Rating: 5.3 |
The non-telic hypothesis for the evolution of life on earth makes no predictions regarding the genome size of the LUCA. However, the non-telic hypothesis does allow for LUCA to have a large genome. Therefore, pointing to a large genome for LUCA does not separate telic from non-telic hypotheses.
But the FLE hypothesis requires that the LUCA have unnecessary but functional genes that would later be used by complex life forms. So does the non-telic hypotheses since it is based on descent with modification.
Non-teleological evolution does not have this requirement; nor does it make any predictions regarding the complexity of the LUCA. What you seem to be missing is that non-telic evolution is about mechanisms, not a specific history. Non-telic evolution is a bottom up process. It starts with simple rules, and from those rules a grander design is produced. When we piece together evolutionary history we look first at the mechanisms and how they are evidenced in the data. The complexity and number of ancestors is a CONCLUSION drawn from the evidence, not a starting assumption or requirement. How evolutionary history played out is a matter of happenstance. What non-telic theories are concerned with are the mechanisms by which these things occurred. Therefore, your claims about the requirements of non-telic hypothesis with respect to LUCA are seriously misplaced. Can non-telic processes produce life that shares deep homology for functional but non-essential genes? Yep, sure can. We have an observed, natural process that is capable of producing the results we see in reality. You want to claim that these same results were instead produced by an unevidenced designer using unevidenced mechanisms. This fails the rule of parsimony, otherwise known as Occam's Razor. The best explanation is the explanation with the fewest unevidenced assumptions. For non-telic processes, the mechanisms are testable and observed. Not so for telic processes.
|
|||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024