
|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| EvC Forum active members: 63 (9162 total) |
|
| |
| popoi | |
| Total: 916,390 Year: 3,647/9,624 Month: 518/974 Week: 131/276 Day: 5/23 Hour: 1/2 |
| Thread ▼ Details |
|
Thread Info
|
|
|
| Author | Topic: Understanding the Baldwin Effect | |||||||||||||||||||||||
|
Ben! Member (Idle past 1419 days)  Posts: 1161 From: Hayward, CA Joined: |
I want to know, how does the Baldwin effect work? My current understanding (developed by reading "Individual Development & Evolution by Gilbert Gottlieb) is that:
Thus, to me, the "Baldwin Effect" is "simply" the development of an "internal trigger" for the development of some morphology that has been selected for. The "internal trigger" removes the dependency on the "environmental change" and simply reuses existing environmental dependencies. How in the world can an "environmental dependency" be incorporated into the organism itself, and become an "internal dependency" (well, really just an "existing environmental dependency" since all development is co-dependent on internal AND external factors)? Note: I did a Google search, and surprisingly at least half of the references I found on the "Baldwin Effect" have to do with computational modelling strategies of behavior. Not far off from my own approach. ("Biological Evolution" ? Or maybe "Education and Creation/Evolution" [after all, this is MY education here < !--UB  --> -->
|
|||||||||||||||||||||||
|
AdminNosy Administrator  Posts: 4754 From: Vancouver, BC, Canada Joined: |
Thread moved here from the Proposed New Topics forum.
|
|||||||||||||||||||||||
|
Ben! Member (Idle past 1419 days) Posts: 1161 From: Hayward, CA Joined: |
|
|||||||||||||||||||||||
|
Saviourmachine Member (Idle past 3574 days)  Posts: 113 From: Holland Joined: |
I have no idea if this falls under the Baldwin effect, but I suppose it has to do with (positive) selection of plasticity.
Natural selection does favour plasticity: organisms that are flexible in adapting to their environment are better in surviving. Genotypes that need a minimal amount of environmental stimuli to develop certain characteristics are more often inherited. The brain is plasticity in optima forma.
|
|||||||||||||||||||||||
|
mick Member (Idle past 5007 days)  Posts: 913 Joined: |
Hi kitanai,
You might have heard recently about the japanese crows which have learned a clever way of opening walnuts. They carry a walnut up onto a traffic light and wait for the lights to go red. When the lights are red, they drop the nut onto the road. They wait patiently while the lights to turn green and the nut is cracked by vehicles. When the lights go red again, they retreive the nut. This behaviour requires a great deal of intelligence, and it seems to be culturally inherited (i.e. crows learn it from each other). The genes that underlie this behaviour are genes that promote learning ability, mimicry, memory, etc. There are lots of genes that will be involved, and they code for plasticity rather than for the specific behaviour. Some birds will be better at learning the behaviour than others. Natural selection will favour genes that enable the behaviour to be learned. For example, a genetic predisposition to remain stationary when they see the colour red is going to be helpful for those dumber birds who aren't able to learn as well as the others. Similarly, a genetic predisposition to cache nuts at a high altitude (i.e. on lampposts or on traffic lights) is going to aid the learning process for those dumber birds. In this way a genetic predisposition toward the behaviour can evolve if the fitness advantage of the learned behaviour is big enough. The behaviour would look the same, but the genetic basis of it would shift from being entirely due to plasticity, to be being partly (or wholly) genetically inherited. I think this is a key point to make about the Baldwin effect. Some morphological or behavioural characteristic might appear in a population due to phenotypic plasticity (i.e. without a direct genetic determination). But natural seleciton might cause the ontological mechanism by which that morphological or behavioural feature arises in an individual to change. The trait itself looks exactly the same. But it arises in an individual by a different mechanism. Hope this helps. It's worth noting that the Baldwin effect hasn't been conclusively proved, as far as I know. It would be difficult, because we would need to identify an evolutionary change in the developmental mechanism that gives rise to the phenotype, and working out these developmental mechanisms for complex traits is very difficult. mick [bold text added in edit] This message has been edited by mick, 03-30-2005 01:10 PM
|
|||||||||||||||||||||||
|
Ben! Member (Idle past 1419 days) Posts: 1161 From: Hayward, CA Joined: |
To me, the Baldwin effect is simply incorporating an external evironmental trigger (i.e. something in excess of the 'normal' environment) for protein transcription into the organism itself, such that the original external trigger becomes unnecessary.
I think this is really different from what you're talking about. BUT, if I can understand how the Baldwin effect ACTUALLY works chemically, then I will be interested to investigate the effects it has on plasticity of the brain. I don't know of any measure of plasticity, but I think if the Baldwin effect were applied to brain-based behavior, I think plasticity would simply be changed (i.e. not increases or decreased) OR decrease.
|
|||||||||||||||||||||||
|
Ben! Member (Idle past 1419 days) Posts: 1161 From: Hayward, CA Joined: |
mick,
Thanks for the reply.
I think this is a key point to make about the Baldwin effect. Some morphological or behavioural characteristic might appear in a population due to phenotypic plasticity (i.e. without a direct genetic determination). After reading "Individual Development & Evolution," I (and the author, Gottlieb) would disagree with you that there's something "without a direct genetic determination." The phenotypic plasticity itself depends on a genetic component. The learning itself also depends on a genetic component. Anything evoking protein transcription is "genetic." As well, there's no such thing as "direct genetic determination." Transcription doesn't happen SANS "an environment." There's always some environment, and there's something in the environment which provides a trigger to transcription. The reason I bring this up is because I think it may help in understanding my overall question. I'm not trying to nit-pick for the sake of it. 
But natural seleciton might cause the ontological mechanism by which that morphological or behavioural feature arises in an individual to change. The trait itself looks exactly the same. But it arises in an individual by a different mechanism. This is a bit different than how I was taking it, but I think there's room for both interpretations of the Baldwin effect. I think the mechanism can be the same, but the "trigger" of the mechanism somehow comes to be internal. My thinking comes directly from Waddington's experiments, where known causes (exposure to a heat shock, exposure to excess amounts of salt) evoked morphological changes in individual development, and selective breeding led to individuals who developed the morphological changes WITHOUT the known cause. In this case, without further information, it seems reasonable (i.e. until further data may show otherwise) to believe that the chemical triggers (original internal chemical trigger brought about by a heat shock, original internal chemical trigger brought about by an excess of salt) that brought about the morphological change are simply triggered by existing protein transcription within the 'normal' development of the organism.
It's worth noting that the Baldwin effect hasn't been conclusively proved, as far as I know. It would be difficult, because we would need to identify an evolutionary change in the developmental mechanism that gives rise to the phenotype Maybe I'm using "the Baldwin effect" poorly. I thought it simply means that observable changes exhibited by individuals only in a changed environment will, if selected for, eventually appear in individuals EVEN IF the original "difference" in the environment is removed. If that's the case, then .. maybe the general case hasn't been proved, but it's certainly been shown to happen experimentally. Even an answer to how those experiments work would be of great interest to me. Thanks again! Edited to fix book author name This message has been edited by Ben, Sunday, 2005/09/18 04:14 PM
|
|||||||||||||||||||||||
|
Saviourmachine Member (Idle past 3574 days) Posts: 113 From: Holland Joined: |
Sorry! You're right.
quote: I failed to notice the second step. It seems a kind of balance thing:
To make my original idea clear:
|
|||||||||||||||||||||||
|
mick Member (Idle past 5007 days) Posts: 913 Joined: |
Ben,
there's no such thing as "direct genetic determination." Okay, I hope this goes without saying. A dominant lethal mutation might appear to be direct genetic determination but obviously it relies on the gene actually being transcribed in a living organism, etc. so I agree even in the reductio absurdium case.
My thinking comes directly from Waddington's experiments, where known causes (exposure to a heat shock, exposure to excess amounts of salt) evoked morphological changes in individual development, and selective breeding led to individuals who developed the morphological changes WITHOUT the known cause I don't think that this is the Baldwin effect precisely. The Baldwin effect is usually applied to social learning. Waddington is talking about "genetic assimilation" or "canalization". His idea is that, under conditions of plasticity within a population, natural selection will act to reinforce the production of whatever phenotype is most useful under the most common environmental conditions. So if morphological changes are evoked by the environment, and if the presence of environmental cues for these changes are consistently present over time, then natural selection will favour genes that give rise to the morphological trait irrespective of whether the environmental cues are present. This is simply because having the trait is the best strategy overall, and producing the trait consistently is a favoured strategy within the population. I'll read up on the Baldwin effect tonight or tomorrow and post back if I can come up with a clear explanation or empirical test. mick
|
|||||||||||||||||||||||
|
Ben! Member (Idle past 1419 days) Posts: 1161 From: Hayward, CA Joined: |
Cool. My understanding comes from Gilbert Gottlieb's discussion on the matter in "Individual Development & Evolution." He discusses two experiments done by Waddington that were used as proof of the Baldwin effect. It is those experiments (giving heat shock to fruitflies prevents some veins in their wings from developing, and the development of some system in their anal papilla when exposed to excessive salt during development) that I am using as the basis of my understanding.
The application to the behavioral domain is more questionable in my mind; I want to understand the original application to mophological entities, to understand how it works (chemically), and then to see if an application to behavior makes sense. Thanks again for taking a look. I appreciate your time. This is really important for the overall approach to understanding behavior that I'm taking. Ben
|
|||||||||||||||||||||||
|
mick Member (Idle past 5007 days) Posts: 913 Joined: |
cheers! Just as a side note, if you are interested in plasticity and evolution, you should definitely read "Developmental Plasticity and Evolution" by Mary Jane West-Eberhard. The amazon link is
http://www.amazon.com/exec/obidos/tg/detail/...{Shorthened display form of URL, to restore page width to normal. - Adminnemooseus} I really can't recommend this book highly enough. And it sounds like its right up your street. I haven't read the Gottlieb book but it's in the library of my Uni so I'll grab it and take a look. Read the West-Eberhard book! Mick This message has been edited by Adminnemooseus, 04-02-2005 12:04 AM
|
|||||||||||||||||||||||
|
Brad McFall Member (Idle past 5053 days)  Posts: 3428 From: Ithaca,NY, USA Joined: |
Waddingtoncross vein paper in Evolution
attempted to relate stabilizing selection to genetic assimilation by opposing it to Schmalhausen's use of "normalizing"as being NOT stabilising quote: and in that interpretation of Wadds experiment it seems possible to try to conceptually link Baldwins' and Waddingtion's notion at the "acquired" character. I however have some reservations from Wadds explanation of his own data but I have not tried to do the most current search on the subject. My Gradfather's data and some discussion on the relation of temperature and wing length in Drosophila is below. 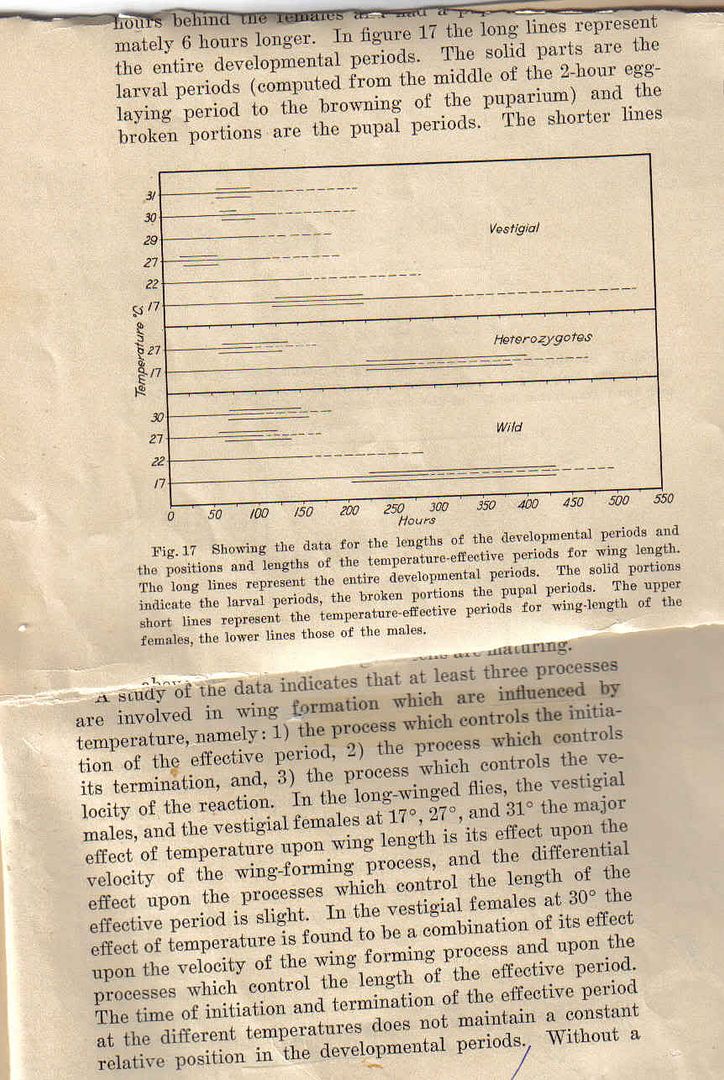 Lets just for the sake say that vestigial being a small wing is like the lack of the cross vein. What Stan did was to experimentally determine the effective period on which different temperatures have effects on chaning the length of the fly wing. He found that the vestigial mutant(think cross veinles mutant) not only reacted to temperature before wild type and heterzygous long wing but the rate of reaction was faster. Thus he figured that three process are involved in the physiology of the effect of temperature on wing size (think cross veinless extent...). Waddingtion's heat shock occurred in the place that are dashed on my Grandfather's graph. Now I give an alternative interpretation. Waddington had said
Discussion
but what if this is NOT "acquired" at all in the sense where Waddington denotes "environment". What if instead the postpupal heat schock simply bumped (on my grandfather's graph) the wild type effective temperature into those for a vestigial (use 30degrees as Stan did not do the study at 40 degrees) but concomittant with this effective period is a lower absolute effective temperature under normal temps than exists for the wild type and thus IS NOT simply the difference of stabilising vs normalizing selection in the subsequent selections "up" and "down".In the experiment described above, selection for the ability to respond to an environmental stimulus has built a strain in which the abnormal phenotype comes to be produced in the absence of any abnormal environment. The crossveinless condition appeared in the foundation stock only as an "acquired character," in the conventional sense. This does not, of course, mean that it was quite independent of the environment; such a supposition would be meaningless, since all development involves not only the genotype but also an appropriate milieu (see Begg, 1952; Waddington, 1952b). What is implied is that crossveinless flies only appeared when the original stock was subjected to some particular abnormal environment, in this case one involving a high temperature at a certain pupal age. In the "high lines" derived from the upward selected stock, however, the crossveinless character appeared in the normal environment which did not involve such a temperature treatment. It has become a hereditary character as contrasted with an acquired one. BulbNrose.com is for sale | HugeDomains Then your interpretation (against) Baldwins'-in favor simply of the chemical "trigger" might be dead on?My grandfather had a quite nuanced proposal for the mechanism of this thermopheneOcopy
here are some potentially interesting links
cross-veinless is an X linked recessive gene.
just a school test the role of phenotypic plasticity in driving evolution discusses crossveinlessquote:This might only be true if the temperature sensitive periods do not prohibit sorting of adaptive peaks. It may be instead that Provine's criticism of the use adapative landscape(gene combinations) in the restricted vision of canalization need not apply to the continuity of the temperature effects through a quantum of kinetic theory of gases. good info Formation of the longitudinal veins (LVs) of the Drosophila wing involves the interplay among Dpp, Egf and Notch pathways. Formation of crossveins (CVs: see Derivatives of the wing disc) present a paradoxical problem. As shown both morphologically and using molecular markers, the definitive CVs are not formed until long after the initial specification of the LVs Such phenotypes are very reminiscent of the crossveinless class of mutations in Drosophila (reviewed in Garcia-Bellido, 1992). This message has been edited by Brad McFall, 04-01-2005 07:56 PM
|
|||||||||||||||||||||||
|
contracycle Inactive Member |
quote: Flip a few variables. Variables can be controlled through either source, and represent either source. matter -> information -> matter ad nauseum
|
|||||||||||||||||||||||
|
Ben! Member (Idle past 1419 days) Posts: 1161 From: Hayward, CA Joined: |
This matches the overall conceptual explanation I was trying to show in the OP. I'm looking for a technical explanation of HOW such a "flipping of variables" would be implemented physically.
I would be greatly indebted if you could offer such a thing. Brad has given such a scenario in the previous post, but sounds like there's probably some more recent information on the subject. Thanks!
|
|||||||||||||||||||||||
|
mick Member (Idle past 5007 days) Posts: 913 Joined: |
Hi Ben,
I did a little more reading and came across three different models of how a morphological trait arising through phenotypic plasticity due to an environmental trigger might become genetically assimilated such that it is produced in later generations in the absence of the environmental trigger. Before explaining them, I want to say that I was wrong when I claimed that the Baldwin effect (and other similar effects) remain to be proven. As you say, they were demonstrated by Waddington and have been repeatedly verified. However there are a few important things to bear in mind, as the effect has only been seen in the lab, under particular conditions. First, it maybe isn’t enough that successive generations of drosophila should suffer a heat shock during early development. This will lead to changes in the frequency of cross-veinless flies, but it won’t necessarily lead to the phenotype being genetically assimilated. Many of these experiments also select individuals displaying the unusual phenotype and only use these to breed the next generation. In these cases, the phenotype appears to be genetically assimilated. So for some of these experiments the assimilation effect may be confounded by the effect of artificial selection. Second, it is worth pointing out that the novel phenotype arising through developmental plasticity is often absent or exceedingly rare in wild-type populations. Waddington’s cross-veinless flies, for example, are rare in nature. In fact the phenotype was produced unexpectedly from the heat shock treatment. It is not thought to confer any adaptive advantage on its bearers. It is just a plastic response that happened to be conveniently produced by heat shock or ether treatment at a very specific developmental stage. Okay, those things out of the way, here are the tree models I could find. They are listed according to how convincing I find them. I should warn you that they are not chemical mechanisms. Mechanisms concerning the interaction of specific gene products simply don’t exist, due to lack of data and theoretical understanding. These are mechanisms that seem more-or-less consistent with our understanding of molecular evolution and gene regulation. There are, however, some computational models of these mechanisms that you can find by searching for something like neural network AND gene regulatory networks on pubmed. 1. Canalization (Waddington) The idea is that there is a typical reaction to some environmental trigger that is directional. For example, when dietary salt is reduced, the anal papillae of drosophila increase in size. There is a straightforward explanation for this — anal papillae are involved in osmoregulation. When dietary salt concentration is low, the cells of the papillae have to increase in size in order to actively transport sufficient salt across the cell membrane. I have no idea whether this hypothesis is correct, Waddington used it in his original articles and I don’t see anybody contradicting it. After a few generations of breeding under low salt conditions, we get the phenotype appearing in the absence of low salt concentrations. The canalization hypothesis states that there has been an increase in the strength of the directional adaptive response, and a partial fixation (or genetic determination) of the response. This implies that the directionality of adaptive responses leads to a directionality of evolution. When Waddington did this experiment, he gradually changed the salt concentration of the food medium such that only 30% of eggs hatched each generation. This is a pretty strong form of selection. The idea is that individuals whose (genetically determined) reaction norm is only weakly directional will be removed from the population, and only individuals with a strongly directional norm will remain. We are selecting for a strong reaction norm. If the norm is so strong that it always produces the required phenotype, then we have what we call genetic assimilation. The important thing about Waddington’s hypothesis is that it is directional. Evolution is more likely, or easier, when it follows the direction of the existing reaction norm. 2. Genetic assimilation in a multigene threshold model (Bateson) Bateson proposed that the plastic phenotype is expressed amongst individuals within a population in a kind of normal distribution. This is also described by Masel (JEB 17(5): 1106. The x-axis of the normal distribution represents the tendency to show the phenotype. The y-axis shows the frequency in the population. The majority of individuals have an intermediate tendency to express the phenotype (the hump of the normal distribution) while some individuals have a very low tendency or a very high tendency. The number of individuals expressing a phenotype under given environmental conditions can be envisaged as the number whose tendency surpasses some environmental threshold. For the heat shock experiment, the cross-veinless phenotype is usually absent in wild-type populations, because while all individuals have some sort of propensity to produce it, the environmental cue does not surpass a basic threshold necessary to activate its expression. The number of individuals expressing the phenotype can be increased either by moving the threshold (i.e. the threshold for expression is reduced by shocking the developing eggs with ether or an abnormal temperature), by increasing the mean tendency for expression (i.e. changing the position of the hump of the normal distribution through natural selection) or by increasing the population variance in the tendency to express the phenotype. Artificial selection under heat shock conditions increases the frequency of cross-veinless alleles in the population, such that a larger proportion of individuals surpass the threshold for appearance of the phenotype in the absence of an environmental cue. The important points are that (a) this model requires that multiple genes to encode the tendency to express the plastic phenotype, and (b) there is no canalization (i.e. no directionality). Masel explains this better than me, it is worth reading his/her paper. I may post the graph, that might be easier. 3. Genetic assimilation under a multigene model with uneven effects (Rendel) The final idea is that a plastic phenotype is based on many different genes at many different loci. I already mentioned that the novel plastic phenotypes that have been shown in drosophila lab experiments are often missing from wild-type populations. The argument is that the uniformity of phenotype in natural populations masks existing genetic heterogeneity for genes involved in the traits production. Imagine that there are ten genes contributing in some way to the veinless phenotype in drosophila. Nine of these have small effects on the phenotype, because of the existence of a single wild-type allele of exceedingly strong effect that counteracts the remainder. Even if all nine alleles contribute to a veinless phenotype, the observed phenotype is veined because of the all important wild-type allele. We might imagine, for example, that the latter suppresses expression of the other nine. Now we bring our drosophila into the lab, and subject them to heat shock. Heat shock interrupts the normal developmental mechanism such that the single dominant wild-type allele no longer exerts its normal influence over the other genes involved in generating veins in the wing. Furthermore, we select flies that show plasticity i.e. those that show an unusual phenotype following heat shock, and use these to breed successive generations. Soon we have lost the wildtype allele, permitting the other alleles to have much greater say in what the final phenotype will become. We can show that the novel phenotype is heritable (because its still based on those other alleles). But the reason it now appears to be heritable whereas it previously only appeared sporadically (or with plasticity) is simply because our experimental procedure removed the dominant wild-type allele from our study population. So we started with a wild type population in which the novel phenotype was absent or rare, removed the wildtype allele from the population, and we now have a different heritable phenotype. Phew! Too much typing. One last warning, that whole lot was based on one evening of reading. I may have grossly misinterpreted these models. I’m sure you or somebody else will let me know if I have! Cheers Mick
|
|||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024








