|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| EvC Forum active members: 65 (9164 total) |
|
| |
| ChatGPT | |
| Total: 916,455 Year: 3,712/9,624 Month: 583/974 Week: 196/276 Day: 36/34 Hour: 2/14 |
| Thread ▼ Details |
|
Member (Idle past 1427 days)  Posts: 20714 From: the other end of the sidewalk Joined: |
|
Thread Info
|
|
|
| Author | Topic: Transitional Fossils Show Evolution in Process | |||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
We often see complaints or comments about the absence of fossil evidence for transitions in evolution. This usually comes in two parts:
(1) There are no transitional fossils
PRATT CC200 quote: Often this is due to a misunderstanding of what "transitional" means in evolutionary biology: Transitional fossil - Wikipedia
quote: Thus all fossils that show intermediate characteristics between ancestral forms and descendant forms are by definition transitional. Thus whenever we see a clear lineage of fossils from an ancestral form (plesiomorphic) to derived descendant form (apomorphic), and thus they are transitional fossils. Transitional fossils will be intermediate in form between ancestral forms and descendant forms, and they will share some, but not all, traits with both ancestors and descendants, and some traits shared by ancestors, the transitional fossil and descendants may themselves be shown in intermediate stages of development, between the ancestral and descendant forms of the traits. (2) There should be billions of transitional fossils
PRATT CC200.1 quote: A recent example of this misunderstanding was presented by Kaichos Man on An ongoing report on S366:Evolution Message 19:
The lack of transitionals that Gould was talking about was small-scale, ie between what would be considered very similar species, neighbours on the tree of life. And this is prcisely what we should see in the fossil record in abundance. Darwin was certain that future fossil finds would support his theory. They didn't. So now neo-Darwinists harp on about fossils being "extremely rare", and "difficult to form". What a load of parrot droppings. Take a look around. Rivers, lakes, seas and oceans everywhere. Daily tides. Frequent droughts and floods. Fossils are being formed by the ton as we speak, all over the world. And this process has been going on for (supposedly) millions of years. If the theory of evolution was true, we would be up to our necks in transitional fossils, each tiny darwinian step lovingly catalogued in the strata. The lack of transitional fossils can only -only- be explained by a lack of transitional species. This thread is intended to discuss and answer this issue. Enjoy. ps - I will also add 3 responses as subthreads, which will cover my initial response at Message 20, so please let me complete those before promotion. Edited by Admin, : Fix rendering. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
In the post that inspired this thread Kaichos Man claimed (Message 19):
The lack of transitional fossils can only -only- be explained by a lack of transitional species. This, of course is the hoary old "absence of evidence is evidence of absence" logical fallacy. The complete absence of fossil evidence for the Coelacanth between the end of the Cretaceous period and modern day clearly proves that these fish did not exist between then and now .... Clearly this statement is false when the Coelacanths are living organisms in the modern world. Other logical fallacies are the argument from ignorance (there are no transitional fossils) and the argument from incredulity (we should be up to our necks in transitional fossils). Logical fallacies are invalid arguments. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
In the post that inspired this thread Kaichos Man claimed (Message 19):
If the theory of evolution was true, we would be up to our necks in transitional fossils, each tiny darwinian step lovingly catalogued in the strata. There are many examples of transitional fossils, and I will discuss two of these as they apply at the species level and they show precisely the "tiny dawinian step" involved in the process of speciation. (1) Foramiinifera:
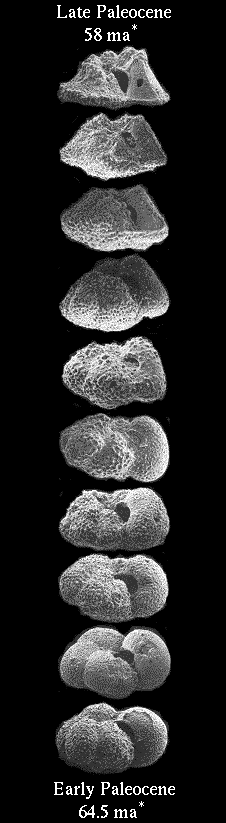
Evolution at SeaComplete Fossil Record from the Ocean Upholds Darwins Gradualism quote: Not just transitional fossils between one species to the next, but the whole pattern of this foraminifera phylum laid out in detail. (2) Pelycodus:
A Smooth Fossil Transition: Pelycodus, a primate quote: This fossil record clearly shows the "tiny dawinian steps" from generation to generation. Note that the "gaps" in time for the fossils are more than covered by the overlap in the variation within each level, each level has organisms similar to the ancestral population below it and to the descendant population above it. Conclusion Clearly transitional fossils exist at the species level, fossils that clearly show the "tiny dawinian steps" from generation to generation. Enjoy. Edited by RAZD, : spling we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
In the post that began this thread Kaichos Man claimed (Message 19):
... this is prcisely what we should see in the fossil record in abundance. Darwin was certain that future fossil finds would support his theory. They didn't. So now neo-Darwinists harp on about fossils being "extremely rare", and "difficult to form". What a load of parrot droppings. Take a look around. Rivers, lakes, seas and oceans everywhere. Daily tides. Frequent droughts and floods. Fossils are being formed by the ton as we speak, all over the world. And this process has been going on for (supposedly) millions of years. If the theory of evolution was true, we would be up to our necks in transitional fossils, each tiny darwinian step lovingly catalogued in the strata. We can ascertain the veracity of this arguement from incredulity by comparing the fossil record for the Foraminifera, Pelycodus and the Coelacanth. As we see in the case of Foraminifera, transitional fossils between species (and higher) exist in abundance at all levels, and in this one case they cover millions of years in a continuous record. This is because the accumulation of these fossils in this location is not reliant on haphazard fossilization, random environmental factors or other things affecting the fossilization of individual organisms. Thus we see, that when there are no causes preventing the reservation of fossils, or for disrupting fossils after deposition, that there is indeed the well preserved record of evolving life year after year, generation after generation, species after species, etc etc, for millions of years. Next we look at Pelycodus and we do see gaps between the fossil layers. There are several reasons such gaps can exist:
Finally, we look at the Coelacanth. The last fossil evidence for Coelacanths is over 65 million years old: Coelacanth - Wikipedia
quote: Here we have a gap in the fossil record of ~65 million years, and yet we have living Coelacanths that clearly show that the absence of fossil evidence is not evidence of absence of Coelacanths. This also shows that fossils do not have to be preserved for intermediate forms. Conclusions What is clear, from comparing these three cases, is:
Simply put, fossils do not need to exist to fill in gaps in the fossil record for intermediate forms to have existed. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi sailorstide, welcome to the fray.
Missing links and or transitional fossils are and can be deemed a non cause and effect for both evolutionary theory and creational theory. Missing links - being missing - don't prove anything other than the evidence is missing. Transitional fossils - fossils intermediate in form between ancestral and descendant forms - can show the hereditary lineage by the progression of change in features over time. This can show the hereditary pattern/s of life from the first forms, whether those are creationist or evolutionary, and thus help us sort between one concept and the other to determine which is the more likely explanation.
I think that both evolutionary and creational theories are just that theories and that when one realizes that both are factually the same thus being theories we can all rest from our own convictions of acceptance. A scientific theory is based on evidence first, then deduced explanation, then testing and then revision as required to explain all the evidence (the scientific process). Thus there is more to a scientific theory than "just a theory" because there is the evidence supporting it, and the testing that validates it and tests it against reality. Evolution fits this definition of theory as a scientific theory. The evidence is the process of evolution we see in the world around us (where all known breeding populations show change in the frequency of hereditary traits from generation to generation), and in the process of speciation (where a parent population divides into two non-interbreeding daughter populations). These are both observed facts. The theory then, is that these two processes can explain the diversity of life as we know it, from the life around us, from history, from archaeology, from the fossil record and from the genetic record. The testing is every new piece of information about past life, whether genetic or fossil, which can either fit the theory or not. Creationism, as far as I can tell, is not founded on evidence, nor does it appear to do any testing of the creationist concept. Rather it seems to be predicated on the a priori assumption that creation is true. This makes it more of an hypothesis, a conjecture, an assumption, rather than a scientific theory based on evidence first, then deduced explanation, then testing and then revision as required to explain all the evidence (the scientific process). But this thread is not about the issue of what is or is not a scientific theory, so if you want to discuss this further, please start a new topic if you want to pursue this, as I'm sure this is a sufficient topic for a new thread. Go to Proposed New Topics to post new topics. I'm sure you will have plenty of takers.
... when one realizes that both are factually the same ... I am not aware of one fact that supports creationism, while I am aware of hundreds that support evolution. Perhaps you can supply us with supporting evidence for creationism? Please start a new topic if you want to pursue this, as I'm sure this is a sufficient topic for a new thread. Go to Proposed New Topics to post new topics. I'm sure you will have plenty of takers.
... we can all rest from our own convictions of acceptance. Unfortunately for you, science is not done by popular vote, it is done by actually confronting the evidence and seeing if the theory passes or fails the tests. Thus when the original post states:
quote: The intent is to discuss the evidence that shows intermediate forms in the fossil record. This is done in Message 3, where clear examples of transitional fossils are presented. The logical conclusions are (a) that transitional fossils exist, (b) they show the same kind of change in hereditary traits in breeding populations from generation to generation as we see in life around us today, and (c) they show the same kind of division of parent populations into non-interbreeding daughter populations as we see in life around us today. They are intermediate. They validate evolution and speciation as being sufficient to explain the diversity seen in the course of their fossil record. Enjoy.
... as you are new here, some posting tips: type [qs]quotes are easy[/qs] and it becomes:
quotes are easy or type [quote]quotes are easy[/quote] and it becomes:
quote: also check out (help) links on any formatting questions when in the reply window. For other formatting tips see Posting Tips If you use the message reply buttons (there's one at the bottom right of each message): ... your message is linked to the one you are replying to (adds clarity). You can also look at the way a post is formatted with the "peek" button next to it. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Taq, love your icon\avatar.
So why do transitional fossils matter in this debate? Because these fossils are a TEST of the theory. How? The theory of evolution predicts which transitionals you should see AND which transitional fossils one should NOT see if the theory is true. This is what is important, how the fossils TEST the theory. The theory predicts that fossils, like modern life, should fall into a nested hierarchy. For example, one should not find a transitional fossil with a mixture of derived mammalian and avian features. Specifically, one should not see a fossil with feathers and three middle ear bones. There are thousands of these types of predictions. A most excellent point, and this also contrasts with the creolution concept of transformation of individual organisms so that there are half formed new features sprouting out of fossils, a cat evolving into a dog, or where new species are formed suddenly by single individuals in one single generation. Creolution claims these are part of "macroevolution", but evolution actually predicts that these type of "transformations" would NOT occur, thus evidence of them would actually be evidence against evolution. Such things are not found in the fossil record or in life around us today. Evolution predicts nested hierarchies via common descent from ancestral populations. In contrast, design (as opposed to creationism ...) would imply that good design features would be replicated, not in a nested hierarchy, but in a manner where new features would be spread across hereditary lineages. We see this repeated again and again in human artifacts and in modern design: the rear windshield wiper appeared on a (iirc) volvo stationwagon, the next year it was seen on many other makes and models, and now is almost ubiquitous on SUVs. We do not see this cross hierarchy design copy pattern in life. Design predicts non-nested hierarchies of features copied across ancestral lineages.
... The main point is that each fossil is a new data point that tests the theory of evolution. Nowhere in the theory does it predict that a fossil from every generation of every species that has ever lived will have been found by the year 2010. However, the theory does predict what mixtures of features these fossils will have when they are found. We now, in the last 50 years, have a second method to verify the nested hierarchy through genetics. Genetics was\is probably the biggest test of evolution, for there is absolutely no reason for a nested hierarchy to appear in the genomes of organisms without common ancestry being true. We see that similar forms occur with convergent evolution, say of sugar gliders and flying squirrels:
So if evolution were not true, that these organisms did not evolve from highly diverse lineages, placental and marsupial diverging long ago, then there should logically be similar DNA for the formation of similar features. Instead genetic analysis says one is placental and the other is marsupial by the nested hierarchies visible in the genetic record. The genetic record confirms the pattern of evolution found in the fossil record. If evolution were not true then there should be homologous DNA for analogous features, and this is not seen in life today.
Gaps are a very distant, secondary concern. It should be noted that gaps do not disprove evolution unless it can be shown that the development of the (theoretical) descendant could not evolve from the last known (proposed) ancestor. These linkages are usually shown as dotted lines of the proposed hierarchy, based on the best morphological evidence available. We are now seeing those gaps being crossed with genetic analysis, where the homologous structure of DNA forms another system of nested hierarchies. Evolution not only predicts that nested hierarchies occur, but that the same nested hierarchies are found in genetics as are found in the fossil record. Like the double helix of DNA, the double pattern of nested hierarchies is seen in life today, entwined one with the other. Enjoy. Edited by RAZD, : copied we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again sailorstide,
Boy some people are really seriuos about their convictions and so am I. Proof and or facts about the existence of GOD and or scientific theories and or ideas are as the words go in the eye of the beholder. Curiously, this has nothing to do with the existence of transitional fossils and whether or not they show evolution in process. Interestingly, there are people that are really serious about talking about the topic of this thread and the factual evidence of reality that exists, whether you believe the evidence or not. Fascinatingly, your opinion has no ability to affect reality in any way. As I noted in reply to your first post, Message 11, this topic is about transitional fossils:
quote: Now if you want to discuss transitional fossils then by all means participate in this thread. However, if you are going to provide us with the depth of your theological\philosophical comments on any other topic then please start a new thread. Go to Proposed New Topics to post new topics.
... I personally believe as a believer in GOD. It may not be written Biblially that this world is 4 billion years old or so yet it is written that this world was here before water, plants and animals were made by GOD and or natural selection. ... Please note that there are many people that are believers in GOD and that have no problem whatsoever reconciling their belief with science in general and evolution in particular. I'm one (see signature). Thus talking about GOD is irrelevant to discussing the reality of transitional fossils, the age of the earth and the geology of the fossil record. So are you going to talk about the topic or start a new thread? Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
AND YOU ARE STILL OFF TOPIC
Edited by RAZD, : subtitle
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Please start a new thread to discuss this.
Sailorsides comments have nada to do with the evidence for transitions in the fossil record. Replying to him only makes the off topic ramble worse.
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Thanks for the reply sailorside,
I was not the one who started the off topic discussion and if you check back on all the replies you will see ... Sorry but the evidence is against you. Your first post was a reply to Briterican, Message 6, and Briterican was on topic, but your reply glanced at the topic and then drifted off into non-topic issues as you stated your personal belief\opinion, and replies to those non-topic issues you introduced went further off-topic, including all of your following replies. Thus if we follow the common descent of replies we find your post Message 7 at the node where they diverge from the topic. But this is relatively irrelevant if we can get back on track and discuss the evidence that transitional fossils show evolution is process. It is extraordinarily easy to diverge from topics when one uses words or phrases that start side discussions so I am not blaming you, just trying to get back on topic.
Is it not so that evidence suggests that all living things have a linking DNA and or RNA make up ? Yes, that is the genetic evidence of common ancestry, evidence that links hereditary lineages in the same way that the morphological evidence in the fossil record links hereditary lineages. This is secondary evidence that the transitional fossils do indeed show evolution in process, as they confirm the pattern of common descent.
Is it not so that all fossils whether 1 million years ago as so indentified and or 100 million years ago as so indentified can have a linking DNA and or RNA link ? Technically yes, however we don't currently have DNA of ancient fossils so this is difficult to confirm for fossils. What we do know is that DNA\RNA evidence shows a pattern of hereditary traits that can be used to develop a pattern of common descent that explains all the evidence. We also know that there is no known reason for this pattern to match and mimic the pattern determined from morphological evaluation of all the fossil evidence, unless common descent is the correct/proper/valid explanation: common descent would predict both patterns. However, genetic evidence is not relevant to the actual pattern of evolution seen in the fossil record - that evidence stands on it's own as a test of the theory of evolution and as noted in Message 11 (my first reply to you):
quote: So we do see validation for common descent and evolution in the transitional fossils that exist in the fossil record. Enjoy. Edited by RAZD, : add Edited by RAZD, : link we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
Hi Kaichos Man - still reading more into information than is there?
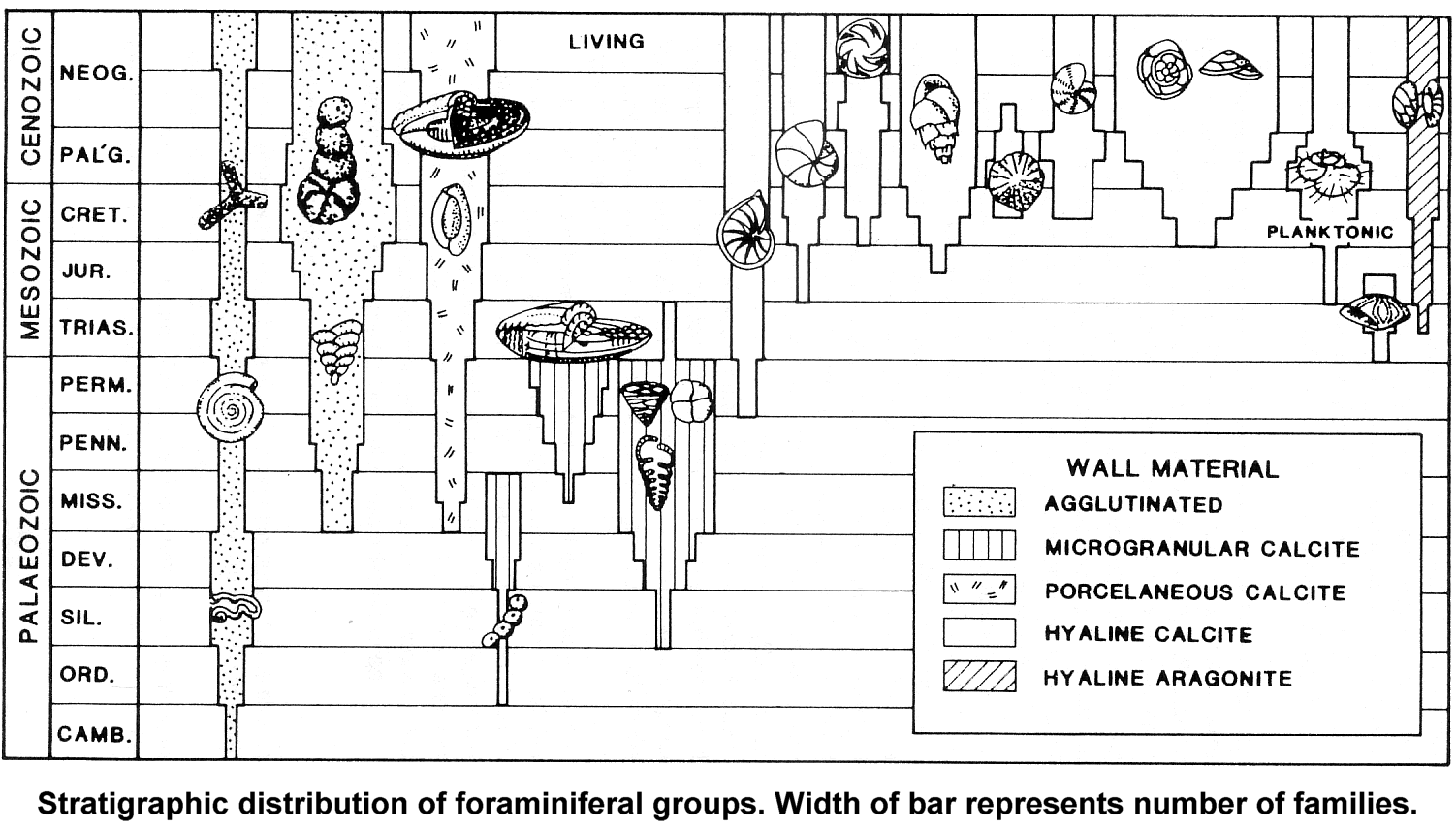
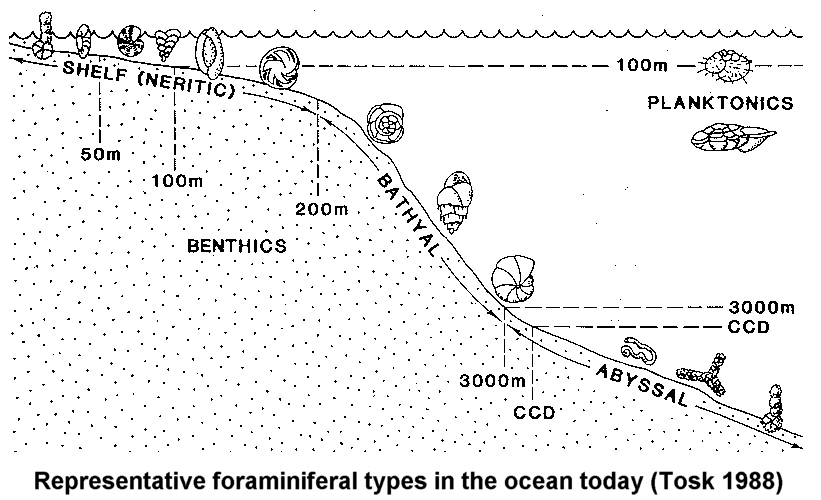
Notice how they differ markedly in relation to their environment. That's because they are ecophynotypic. That should have sounded a warning bell for Tony Arnold and Bill Parker but it obviously didn't. However, fortunately there are more professional scientists around: There are a couple of problems here. Notice at the right edge of your graphic there is a group labeled "planktonic" and the rest are all benthic. Your article applies to benthic forms:
quote: While Arnold and Parker studied planktonic:
Evolution at Sea quote: So the free floating ones are not bound by the ecological constraints of their local environment the way the benthic ones are. Furthermore, your article only covers two groups of benthic forams: http://www.springerlink.com/content/83502273g54060w5
quote: However, similar studies have been done on the planktonic forams with similar results of finding cryptic species. Foraminifera - Wikipedia
quote: Where "morphospecies" are species groups defined by their morphology, while understanding that there may be one or more cryptic genetic species involved. This means there are possibly more species, but it is difficult to say because they look the same. So finally, there is the problem of what the evidence actually shows, versus what you claimed:
... They underestimated diversity. They got it wrong. They thought they were looking at different critters, when they were the same critters wearing different coats. Ecophenotypes, RAZD. Not evolutionary successors. ... You have it exactly backwards -- the cryptic forams were wearing the "same coats" while exhibiting greater diversity under those "coats" (tests). At worst this means that where Arnold and Parker saw some speciation events, they may have missed others due to crypsis, and this in no way invalidates the speciation events seen, nor does it invalidate the panorama of transitional forms for the planktonic forams. Classifying forams by their tests ("coats") underestimates the diversity, but does not invalidate the fossil record of change in breeding population from generation to generation. Enjoy we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Further to the previous reply, the article cited can be found here:
Cryptic species of planktonic foraminifera: thier effect on paleoceanographic reconstructions, by Kucera, M., and Darling, K.F., 2002. Bits and pieces from the article:
quote: Things to note:
None of this recent work invalidates the fossil record showing lineages of common descent with changes in morphology over time, nor does it invalidate the observed division of parent populations into reproductively isolated daughter populations, rather this new work reinforces this pattern and extends it to a finer detail in the living species. This same level of detail may not be discernible in the fossil record, yet we can assume that it exists or not, and the pattern of common descent still holds, locked in the fossils, evidence of intermediate forms between ancestral and descendants, evidence of evolution in process: forams are indeed transitional fossils, as the term is used in science. Enjoy. Edited by RAZD, : more links Edited by RAZD, : glitch fixed we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
Hi Kaichos Man, you will be pleased to know that I gave your post serious consideration. I'm going to go long on you, because you deserve it.
When you first posted the question about ecophenotypes I noticed that you provided no evidence to link the term to foraminifera other than your claim. Previous experience with your claims leaves this a questionable source of authority at best.
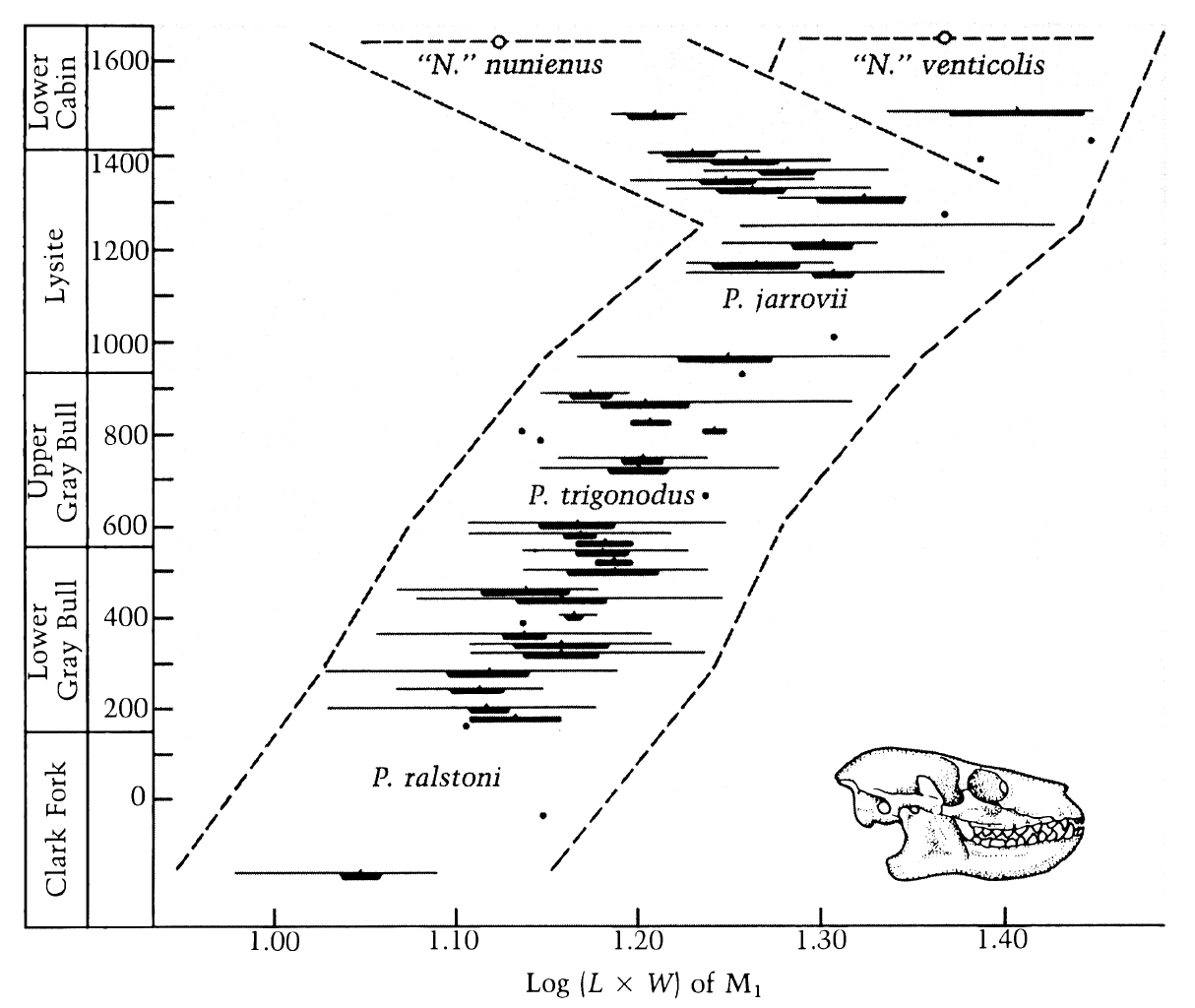
Message 30: Notice how they differ markedly in relation to their environment. That's because they are ecophynotypic. It seemed fairly obvious to me that you had, once again, willfully misinterpreted some piece of information. I notice you've changed the picture - the other one showed better the divisions of the different groups of foraminifera (remember that this is a phylum not a species), however I can still work with this new graph as it shows the major groupings of forams. There are ~13 known orders of foraminifera, with more taxon divisions below that. Parker and Arnold said that they had documented over 300 species ("Counting both living and extinct animals, about 330 species of planktonic forams have been classified so far, Arnold said."), so there are likely quite a number of extant species. Here is your previous image for reference (from your actual source):
quote: Notice that the width of the bars in this graphic represent the number of families within each group, so we still are not down to the species level or even the genus level. BTW -- I'll echo Percy here: if you are going to post pictures or quote sections of articles you should provide links to your sources as part of your evidence, it's that old thing about proper credit where it is due eh? Here is the text that accompanies your current picture:
quote: Evidently Tosk is a creationist trying desperately to explain away the vast geological ages and massive data on foraminifera with a global flood and half vast imagination. You should use (a) more current resources and (b) more reliable resources. Let's continue: This latest picture shows 14 different examples of forams occupying different ecologies, and this may be where the known orders were when it was published (in 1988). Certainly we cannot assume that this picture represents species or genera or even families of forams, as too few are shown. The second reason I thought you were blowing smoke, was that I had not run across the term you gave in any previous reading on forams, and it was not mentioned in the article on forams in wikipedia (not that this is an authority, just a relatively current referential starting point). The third reason I thought you had it all wrong, was that the article you quoted (without source) contradicted what you said:
quote: Percy found the link to the abstract you quoted from:
Message 33: Here's a link to the article by Pawlowski and Holzmann whose abstract you quoted from:
Where does it say anything supporting your claim that Arnold and Parker confused morphological differences with species differences? And here is the whole abstract:
quote: There are a couple of things to note here:
Now, in general, when biologists talk about variations within a species they use the term variety. Several different varieties can exist within a single species, and it is common in many species to have distinctive varieties. To have ecophenotypic variants you would need to have distinctly different varieties within a genetic species. Conversely, when biologists generally talk about differences between species they talk about diversity, when speciation occurs the parent population diversifies into two distinct species. Cryptic means that different species look alike: Species complex - Wikipedia
quote: ... italics for emphasis. So when the paper says that both examples "perfectly illustrate high cryptic diversity revealed in almost all molecular studies" they specifically mean that there are cryptic species that look very similar but that they are genetically distinct. They may be (likely are) closely related (especially given that Ammonia is a genus), but they are not variations within a single species. Likewise when they say that "morphology-based studies largely underestimated foraminiferal diversity" they means that there are more species than is readily apparent from just looking at the morphology due to the cryptic species looking so similar. Entirely the opposite of what your creationist website tries to pretend. Now I though I made this point clear when I posted the quote from the wikipedia article on forams (that mentions morphospecies but does not mention ecophenotypes):
Message 34 Foraminifera - Wikipedia quote: Where "morphospecies" are species groups defined by their morphology, while understanding that there may be one or more cryptic genetic species involved. This means there are possibly more species, but it is difficult to say because they look the same. Notice that when we talk about modern planktonic forams, that there are some 40 morphospecies, 40 groups that are morphologically different. Now I though I drove this point home in the next post when I provided you with a second reference, this one on planktonic foraminifera, similar to the ones studied by Parker and Arnold, that ALSO talked about morphospecies:
Message 36 (edited for brevity): Further to the previous reply, the article cited can be found here:
Cryptic species of planktonic foraminifera: thier effect on paleoceanographic reconstructions, by Kucera, M., and Darling, K.F., 2002. Bits and pieces from the article:
quote: Things to note:
Notice that all the references to classification of forams as "ecophenotypic variants" are dated 1976 or earlier. Before Parker and Arnold (so they would be aware of this possibility) and before DNA sequencing for genetic analysis. Notice that this paper shows that what appeared to be "ecophenotypic variants" is now, by genetic analysis, seen as the genetic variation within the different morphospecies explained by the different genetic types, species classifications based on DNA instead. Now we come to your recent post.
I bring up "ecophenotypes", a single species exhibiting a range of morphotypes. Then you bring up "morphospecies", which can be several species exhibiting the same morphotype- and accuse me of having it the wrong way around! First, you will note that I did not introduce morphospecies, I provided the evidence from scientific studies of actual forams by scientists who classified them as morphospecies, and went on to show that the articles were indeed talking about morphospecies and not ecophenotypes. These articles show that you had it backwards.
The fact is, forams are extraordinarily plastic. They possess, as you have pointed out, "cryptic genetic variation". It may be worthwhile to establish exactly what that means: And again, you provide no reference of actual science done on forams to substantiate this claim. Please note that nowhere in my post did I mention "cryptic genetic variation" -- that this is YOUR misrepresentation of the argument against you. I mentioned cryptic species (because the articles talk about cryptic species), and I mentioned cryptic genetic diversity (reference above to terminology).
The cryptic genetic diversity of living foraminifera has been established through molecular studies. But the forams comprising the fossil record contain no DNA. That's why Arnold and Parker had to rely on microphotography. So I went looking for scientific articles on forams and ecophenotypes to see what I could find.
quote: Note (1) that this is 1978, and (2) that these forams involved ("Ammotium, Ammonia, and Elphidium") are three benthic genera, one of which Ammonia is specifically referred to in the article on benthic forams that showed cryptic genetic diversity instead of ecophenotypic variation. New information displaces old mistakes.
quote: 1986 - still old stuff, and ... Ammonia again. The other articles I found mentioned ecophenotypes because they cited the old papers as part of the history of classifying forams. One of these papers is the one I've provided in Message 36. By the time that Parker and Arnold made their morphological analysis of all the known marine planktonic forams, the idea of ecophenotypes had pretty much disappeared, and when you get to all the current articles that are based on genetic analysis we see that previous classification involving "ecophenotypic variants" were eroneous, and that the slight morphological differences were due to real genetic differences between cryptic species.
It means that when you look at three morphotypes you could be looking at six species or one. And the fact is, it's impossible to tell: No, Kaichos Man, it is not always possible to tell when you just look at the morphologies, however when you use genetic analysis and determine that there are distinct different genotypes involved you can tell, due to real genetic differences between cryptic species. This is what science has done since 1988. It has invalidated your premise that ecophenotypes are rampant through the phylum, and it has consistently shown that there are more genetic species instead -- more diversity, not less. Parker and Arnold showed what a robust morphological analysis could do to the biogeology of forams, but they underestimated the diversity of species involved. As such their morphological tree of descent from common ancestors stands, not dismembered, but stronger as it is validated by the genetic analysis of planktonic forams. The transitions and speciation events they show are still valid transitional fossils, although they may represent genera instead of species. Enjoy. ps -- thanks with providing me with another creationist hoax site (for Message 56):The Emperor Has No Clothes - Naturalism and The Theory of Evolution telling lies to gullible believers Edited by RAZD, : link Edited by RAZD, : clrty we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Well Kaichos Man, what do you think?
You are suggesting then, RAZD, that ecophenotypic variation doesn't occur in foraminifera? The evidence shows that whenever genetic analysis is done, that no evidence for ecophenotypic variation is found, and in it's place, several cryptic species are found that are more than adequate to explain the previous old (1976) idea that ecophenotypic variation was involved. The evidence shows that the text accompanying your second picture, the one by creationist Trosk, is false, and that the different orders of forams are not all one species. His diagram is obviously a depiction of many of the same shells as the first diagram, which represents the orders of forams, not species, and when species are genetically distinct within the genus level, any attempt to claim that orders are one species is just plain ridiculous. Trosk is a discredited charlatan, and "Sean D. Pitman M.D." is either a gullible fool, delusional, ignorant of reality, or intentionally lying (your choice). You have the opportunity to learn from his mistake. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1427 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again hawkes nightmare, finding your way around?
ok, about those pictures with the sugar glider and the flying squirrel. ... You mean the example of convergent evolution that was given in Message 14:
We now, in the last 50 years, have a second method to verify the nested hierarchy through genetics. Genetics was\is probably the biggest test of evolution, for there is absolutely no reason for a nested hierarchy to appear in the genomes of organisms without common ancestry being true. We see that similar forms occur with convergent evolution, say of sugar gliders and flying squirrels:
So if evolution were not true, that these organisms did not evolve from highly diverse lineages, placental and marsupial diverging long ago, then there should logically be similar DNA for the formation of similar features. Instead genetic analysis says one is placental and the other is marsupial by the nested hierarchies visible in the genetic record. The genetic record confirms the pattern of evolution found in the fossil record. If evolution were not true then there should be homologous DNA for analogous features, and this is not seen in life today. If you reply to the message with the information you are replying to with the message reply button (there's one at the bottom right of each message):... your message is linked to the one you are replying to (adds clarity). This helps people track back to see what the previous post was about. You can also look at the way a post is formatted with the "peek" button next to it.
... what did the prehistoric sugar gliders look like? where are the transitional fossils between where they didn't have extra skin, to the ones where the started to develop it, to the present picture? show me. Why? You will note that these cute little guys are not presented as intermediate fossils, but rather part of a secondary test of evolutionary theory. The fact is that skin rarely fossilizes, and so such features are hard to distinguish in the fossil record -- they may be there but not noticed as ancestral to the sugar glider or flying squirrel. What we do know from the skeletal and genetic evidence of both animals, is that they have distinctively different features and traits under the skin, and that the appearance of similarity is just that: superficial skin deep appearance. The appearance of similar design would argue that similar DNA would be involved for the development of each, but this is not the case. This is an opportunity, however, to introduce another transitional fossil:
quote: Note that there is no fossil evidence of the skin, so the attachment of the skin to the skeleton is inferred from secondary evidence. This is the abstract link for the Nature article (you will need sign in privilege to read the full text):
quote: Here is the graphic showing the plot of limb ratios from the full article in Nature:
You will notice that Onychonycteris finneyi is exactly between the non-flying cohorts and the flying bats known in the fossil record and modern day. Note that Symphalangus, the blue triangle closest to this fossil, is a gibbon. Bradypodidae, the next closest, are sloths, Sciuridae, near the bottom left, are squirrels, and this would include the flying squirrel. Scandentia are tree shrews, thought by many to share a common ancestor with bats. Cynocephalus is a "flying" lemur:
A transitional fossil is one that shows traits intermediate between ancestral forms and descendant forms, and this bat is clearly between modern bats and non-flying arboreal organisms. Enjoy.
... as you are new here, some posting tips: type [qs]quotes are easy[/qs] and it becomes:
quotes are easy or type [quote]quotes are easy[/quote] and it becomes:
quote: also check out (help) links on any formatting questions when in the reply window. For other formatting tips see Posting Tips Edited by RAZD, : addd we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024
 (1)
(1)