|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| EvC Forum active members: 64 (9164 total) |
|
| |
| ChatGPT | |
| Total: 916,815 Year: 4,072/9,624 Month: 943/974 Week: 270/286 Day: 31/46 Hour: 3/3 |
| Thread ▼ Details |
|
Member (Idle past 1432 days)  Posts: 20714 From: the other end of the sidewalk Joined: |
|
Thread Info
|
|
|
| Author | Topic: Transitional Fossils Show Evolution in Process | |||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again hawkes nightmare,
but i believe it was just a type of bat that died out because it couldn't adapt o its environment, not a transitional species You are welcome to your opinion, however opinion is curiously incapable of altering reality to match.
but i know the layout of forums, as i am currently registered to about 5 or 6 of them. And this forum has some features not found on many. This of course, is due to intelligent design, but that's a different topic for a different thread. Enjoy we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi hawkes nightmare.
here's you disproof: Bombardier Beetles and the Argument of Design If you want to discuss this PRATT please start a new thread. Go to Proposed New Topics to post new topics. One of the things we like to do in the forum is stay on topic. This has nothing to do with transitional fossils. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
Just to be clear, hawkes nightmare,
no i shouldn't because it is relevant to this thread. cyote said he wanted animals that disprove evolution, i gave him three websites Coyote's comment is not the topic. The topic is transitional fossils show evolution in process. Read Message 1: that is the topic. If you want to discuss your websites with Coyote, he is waiting for you here
PRATT Party and Free for All, Message 1. Note also that bare links with no comment are violations of forum guidelines. You may want to read them to be sure they are like other forums:
Forum Guidelines Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Still grasping at hope, Kaichos Man?
I understand why you have embarked on this Quixotic mission to destroy the concept of ecophenotypy, RAZD. You appreciate that it renders the foraminiferal fossil record nonsensical as "an unbroken evolutionary progression". Curiously it doesn't. All the articles that mention ecophenotypes are discussing species varieties already known at the time, and in no case that I have seen has anyone said that two species from outside of a genus were found to be ecophenotypic varieties of a single species. Here's the 1978 paper by Helen Tappan:
quote: Notice that she talks about variations within a species that are already known as variants, and she explains it by ecophenotypic variation. Notice that she also lists two such species from the same genus, Elphidium but does not suggest that they are one species. Notice that one of her ecophenotype species, Ammonia parkinsoniana comes from the genus that was later found to be composed of cryptic species when genetic analysis was done. Here's another article:
quote: So we have another species with known ecophenotype variants, and we still see change in the frequency of traits in their populations from generation to generation, either in response to environmental factors, or to adaptation within the ecophenotypes for different ecological preferences. Looks like that still supports the topic thesis that Transitional Fossils Show Evolution in Process. This is the best case for ecophenotypes I could find, and it is not any comfort for your position, because it does not change any species classifications or group any species together into a new super species, and that is the kind of result you need to invalidate Parker and Arnold. Note that I did find this in google:
quote: Unfortunately the abstract says nothing more about Parker, but we can certainly feel confident that he was well aware of the issue of ecophenotypic variation in fossil foraminifera when he and Arnold made their morphological analysis. Here's the abstract:
quote: In other words, the people developing the automated image analysis systems, such as the one later used by Parker and Arnold to analyze all known planktonic foraminifera, were calibrating them against known instances of ecophenotypic variations within certain known species. With this calibration the system then identifies the variations within Neogloboquadrina pachyderma and Globorotalia inflata as being members of a single species, and not divided out into new species based on morphology alone. The result is a conservative underestimation of species diversity, where some differences in morphology may be grouped together when they are actually distinct species, as has been found to be the case when genetic analysis is done. This further weakens your position. And your link doesn't necessarily prove your case either, only that they are discussing whether it is one or the other when they say (see italics):
quote: Want to provide the link so we can see what the whole abstract says? I could not find it, however I did find this:
quote: Again, it looks like division into subspecies (ie a subpopulation variety) is more likely than a single species with different development in different ecologies. It will be interesting to see what happens when we get to genetic analysis eh? I'd bet on sibling cryptospecies within a morphospecies resulting in increased diversity one more time, but it isn't necessary. Do you understand that whether they are ecophenotypic varieties within a single known species, or actually a morphospecies with multiple cryptic species, that the classification is still a group of forams that have evolved as different genetic lineages from other foram species? Do you understand that this does not affect the tree of life of common ancestry as determined by Parker and Arnold? Do you understand that they are not saying that Uvigerina peregrina is the same species as Globorotalia truncatulinoides, Ammonia parkinsoniana, Elphidium gunteri, Elphidium galvestonense, Palmerinella palmerae, AND Ammotium salsum - as was falsely claimed by Troste and Pitman? That would be the kind of information you need, not articles discussing the level of variation seen within these species as they are currently classified.
Try googling "ecophenotypic foraminifera", and you'll see what you're up against. I have, and I have found very few articles (per information above) that actually talk about ecophenotypes existing, rather than articles where ecophenotypes are being reclassified as cryptic species or distinct subpopulation varieties. I also went to the home page for The Journal of Foraminal Research:http://jfr.geoscienceworld.org/ and a search of their data base for "ecophenotype" turned up three (3) articles: quote: The first one is already listed above, the second talks about Ammonia, Tritaxis and Elphidium, two of which have already been discussed, and the abstract for the third article mentions neither ecophenotypes nor morphospecies. This is three out of hundreds and hundreds of articles about foraminifera.
Some heavy names there, RAZD. And they obviously see ecophenotypes not only as a possibility but as a real problem in using this particular foram as a proxy. And again, your conclusion just does not follow from the data: they are not regrouping species together into a new superspecies, but discussing existing classifications, and in many cases refining the definitions with either sybling cryptic species or subspecies populations. As for your "heavy names" attempted argument from authority, I note that Jan Pawlowski is one of the authors of the article we've discussed previously (Message 40):
And here is the whole abstract:
quote:... So when the paper says that both examples "perfectly illustrate high cryptic diversity revealed in almost all molecular studies" they specifically mean that there are cryptic species that look very similar but that they are genetically distinct. ... Likewise when they say that "morphology-based studies largely underestimated foraminiferal diversity" they means that there are more species than is readily apparent from just looking at the morphology due to the cryptic species looking so similar. Entirely the opposite of what your creationist website tries to pretend. So I'm still not finding any evidence that begins to support your position.
But how on earth are you going to counter all of that scientific literature? Try googling "ecophenotypic foraminifera", and you'll see what you're up against. Curiously, that is how I found all the articles on morphospecies and cryptic species. For instance, here is another one by those "heavy names" Jan Pawlowsli and Maria Holzmann:
quote: We still see genetic analysis finding cryptic species replacing previous classifications of ecophenotypes. We still find that
"The evidence shows that whenever genetic analysis is done, that no evidence for ecophenotypic variation is found, and in it's place, several cryptic species are found that are more than adequate to explain the previous old (1976) idea that ecophenotypic variation was involved." That's a pretty sweeping statement. Let's see if the latest literature by the top scientists supports it. If it is wrong, then it should be a simple matter for you to find a single article that refutes it. The fact is, still, that none of your articles yet posted either support your position or invalidates that statement. It seems that the issue of ecophenotypic variants is already known in the study of forams, and that the degree of variation involved is not enough to turn the phylum of foraminifera into a single species. If anything, this has led to a conservative classification of foram species into general morphological groupings that includes the ecophenotypic variation, and that rather than the overestimating the numbers of species this has underestimated the actual species diversity. Nor is the degree of variation involved sufficient to invalidate the hundreds of speciation events and evolutionary lineages of descent found in the fossil record, as found by Parker and Arnold. When we see transitions like this in the fossil record:
Transitional Fossils Show Evolution in Process. Enjoy. Edited by RAZD, : clrty we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Percy,
Google scholar gets 104,000 hits for "foraminifera" and only 456 for "foraminifera ecophenotypic". It's worse than that: when you google ecophenotype foraminifera, and this thread on EvC is the second hit. The first hit is Helen Tappan's 1978 paper. ecophenotype foraminifera - Google Search Conclusion: your forum is one of the leading experts on ecophenotype foraminifera ... or it just is not that big an issue. Enjoy. Edited by RAZD, : link we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again, Kaichos Man, let's see if we can clear some things up.
Do you understand that whether they are ecophenotypic varieties within a single known species, or actually a morphospecies with multiple cryptic species, that the classification is still a group of forams that have evolved as different genetic lineages from other foram species?
Yes I do. So classificiations of the forams at the genus level is valid regardless of whether we have ecophenotype variants or subspecies population variants. Consider this the fall back and punt position for the tree of life as determined by Parker and Arnold, and this should make the next point easier.
Do you understand that this does not affect the tree of life of common ancestry as determined by Parker and Arnold?
No I don't. The first branch in on the tree of life for any species is to their genus category, so if the genus classifications are all valid, then the arrangement from genus down to the first foram population would be valid. Note references in [brackets] below are to abstracts and quotes provided in Message 58. We may see some additional classification of species within morphospecies to account for the ("underestimated") genetic species diversity that is being discovered through genetic analysis: this adds branches to the tree of life (to incorporate the "cryptic genetic types" [Kucera and Darling, 2002]), with the morpospecies as a likely common ancestor (an example of why classical taxonomy gets messy compared to cladogenetic classification). We may see some additional subdivision of species into subspecies or variety classifications ("Uvigerina peregrina parva is most likely a subspecies of Uvigerina peregrina and not an ecophenotype" [Schnfeld, 2006]): these would be stable subpopulations with varietal traits fixed in their genes, but still be part of the biological species in question (like the varieties of asian greenish warblers or the varieties of peppered moths). We may see some confirmation of ecophenotypic variants within some species ("Field and laboratory evidence demonstrates that this paired ecophenotypy is caused by contrasting results of delayed reproductive maturation in minimum environments, versus accelerated maturation in optimum environments. Longer growth periods produce larger, thickly calcified tests; shorter growth periods produce smaller, thinly calcified tests." [Tappan, 1978]). Such variation may well be due to aspects of the water column ("Analysis of the planktonic foraminiferal species, Neogloboquadrina pachyderma and Globorotalia inflata, indicate that the test outlines of these species reflect various physical parameters of the water column." [Healy-Williams, 2007]). Note that my impression from reading all these articles is that the known ecophenotypic variation is relatively small and apparently more a matter of maturation than different development, of overall complexity rather than markedly different pattern/s in the tests, ie - variation between single chamber and multichamber, but not to different multichamber patterns. The proposed ecological causes are pressure, salinity and levels of nutrition are plausible for such changes. With the advent of genetic analysis, it is possible that some species will be reclassified from one genus to another, but this will not be due to their being ecophenotypes, it will be due to evidence of genetic heredity and cladistic analysis. It is highly unlikely, however, that any substantive change to current arrangement of the tree of life for planktonic (or benthic) foraminifera will be made, as the classification of the various genera has not (yet) been challenged by any new findings since Parker and Arnold's work. What we are likely to see is refinement of it through additional information.
Regarding Arnold and Palmer's evolutionary progression as real science is like regarding Hans Christian Anderson's work as a factual history of Denmark. And yet neither you nor any article yet found has shown how a single aspect of Parker and Arnold's morphological analysis is invalid, even at the species level. (Note - I kept wanting to type Palmer too, btw, so I reversed the order to make it easier to remember properly). enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again Kaichos Man
You miss the point, RAZD. I am not putting forward an unbroken evolutionary progression. The onus is not on me to show it is invalid. It is on Parker and Arnold to prove that it is valid. No, sorry. If you think their conclusion is false you need to show evidence that invalidates it, not just wish that it is invalid. Science works by invalidation of false concepts, not by proofs. If you don't understand this, then you don't understand either logic or science.
Let's take another look at Dr Michael Knappertsbusch's paper:
quote: First off, I'll take this as a tacit admission that your first issue with ecophenotypic variation does not have the traction you need to disrupt Parker and Arnold's work, as now you are changing horses in typical creationist manner. Again, you did not provide a link to your reference, but I've found it anyway.
quote: Note (1) that all four of these varieties are within the genus Globorotalia and that all are direct descendants from the Globorotalia menardii species:
... so as noted in Message 62:
quote: So this still does not challenge the work of Parker and Arnold in any way. Note (2) that the variations noted are variations on a theme displayed during the development of the forams with minor variation in complexity:
"... Measurements were made of spire height (∂x), maximum diameter (∂y), the tangent angles of the upper and lower peripheral keels (Φ1, Φ2, respectively), the number of chambers in the final whorl, and the area of the silhouette in keel view. Four morphotypes alpha, beta, gamma, and delta were distinguished. ... " ... and as such show even less variation than I said (Message 62) may be expected:
quote: So nothing here challenges Parker and Arnold. The whole paper is available at this link, rather than just the abstract, so we can look at it in greater depth, rather than just extract a tid-bit or two that fits your confirmation bias and ill/uninformed opinion:
Let's take another look at Dr Michael Knappertsbusch's paper:
quote: Now let's read the preceding paragraph:
quote: One such study was the one by Parker and Arnold eh? We also saw reference in Message 58 to similar work done by Healy-Williams (2007). Note that this is a "powerful means of studying speciation in the geological past," and not one that is being criticized, discarded, or found wanting. Your three quotes come in the next paragraph:
quote: But it doesn't stop there: now let's look at what he says after your quoted tid-bit:
quote: In other words, this "major difficulty" can be overcome by three different methodologies. Let's see what his conclusions are:
quote: Again, the amount and type of variation exhibited by Globorotalia menardii and its descendants is much less than the differences between the diverse and wonderous forms of the different genera of forams. Here's a few pictures of these four forms:
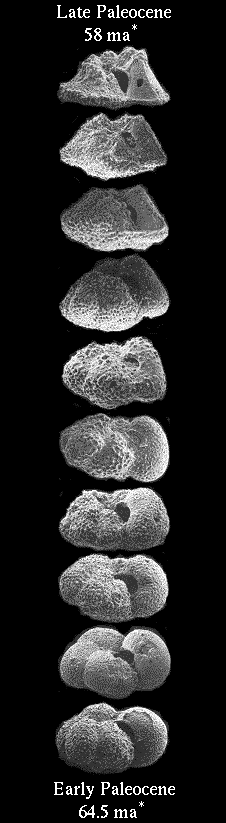
Compare those differences with the amount of difference shown in the Parker and Arnold sequence here:
Does that look like earth shattering morphological differences that challenge the classification of foraminifera on the basis of morphological analysis as done by Parker and Arnold? Looks like my comments in Message 62 still stand unchallenged:
quote: Indeed, this article is validation that what we are seeing is refinement through additional information that builds on the previous morphological studies rather than invalidate them. Enjoy. Edited by RAZD, : clrty Edited by RAZD, : comparison added we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
It is a typical creationist excuse to claim partisan bias, Kaichos Man, especially when there is none.
... on that clearly partisan note, I shall quit the forum. The problem is that there is no partisan requirement here. A fact Percy noted when he said:
Because this is an objective criteria I will continue as a participant in the discussion. You were not asked to do anything I have not done, and you were in violation of the Forum Rules quote: The reason is more than just being able to verify that your quotes are accurate representations of the material, it is basic honesty and part of intellectual integrity. More likely, this is just an excuse you tell yourself to avoid dealing with the issues that show your argument is weak at best, and false at worse, as you have been completely unable to justify your claims in any way. A typical ploy when a creationist loses an argument is to declare victory (usually on some "moral" grounds) and run away. Enjoy. Edited by RAZD, : 6 we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again Kaichos Man
Message 73: Nowhere that I can see in the guidelines is an actual link to the source required. However, if this has become a new requirement, I am happy to abide by it. It's rather implicit in rule 6:
quote: Message 74: And who might you be? I've posted 189 times, and the only one you replied to was the one where you thought I wasn't coming back. There are lots of people that read these threads even thought they don't reply. I often don't reply to threads when other people are saying the things I would say, as that would only clutter up the thread and overload posters with many replies to the same post. I'm sure you've experienced this.
You see a significant difference between the terms "ecophenotypic variation" and "clinal morphology"? Yes. The time scale is different and the type of adaptation is different. Ecotypic variation occurs during the development of individual organisms in response to ecological factors, like pressure, salinity, temperature, average amounts of light, food availability, etc etc etc. In a different ecology they would develop differently. Whereas "clinal morphological changes due to coadaptation to similar environmental gradients" occur over generations and involve genetic adaptation: Cline (biology) - Wikipedia
quote: Thus clinal morphological variation is similar to subspecies population variation, except that the populations overlap. In a different ecology they would (and do - hence the overlap) still develop the same.
Your impression? Honestly, RAZD, has it come down to that? We've got Percy gauging veracity by the number of mentions on Google Scholar, and you putting forward your impressions. Well, Kaichos Man, you certainly have not provided any information that would show otherwise. You've raised a point you think is a telling blow to the work of Parker and Arnold, but nobody can find evidence that shows more than minor variations within species in your references (when we can find them) or outright lies (like your creationist site).
So having referenced (indirectly) the work of Parker and Arnold, Dr Knappertsbush then raised a "major difficulty" in foraminiferal taxonomy and goes on to list three possible solutions- none of which applies to the work of Parker and Arnold. Right, none of which affects the work of Parker and Arnold because it is all about the minor variations within species, with at most a mix up of closely related species (each directly descended from the same parent species). Look again at what defines the "major problem":
quote: In otherwords, the difficulty is in distinguishing species differentiation from subpopulation variation in clines from ecophenotype variation in individuals. It seems that the "major difficulty" is sorting out subspecies versus species, a rather common problem in taxonomy (see Asian Greenish Warblers as an example of this problem). This still does not affect the overall lineages of descent from parent populations and the changes in species seen over millions of years as sorted out by Parker and Arnold, as the only effect is whether the ends have one or several twigs at the tips. Enjoy. Edited by RAZD, : more we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Foxdog and welcome to the fray.
In practice, Evolution is neither fact, nor theory ... A few quick points on distinguishing the three aspects of evolution:
Contrary to the assertions of main stream evolutionists (who also just happen to be atheists by the way), ... Why do creationists keep making this easily falsified assertion? I'm no atheist, and there are more theist evolutionists than atheists.
Sure one can assert that evolution is observable through fossil records; but if the records were as clear and unambiguous as many claim then there wouldnt be any disagreement among scientists. And yet there is very little disagreement on the validity of evolution - how can that be ... unless the disagreements are about minor points rather than the major structure? If you want to discuss these issues further, please start a new thread.
The problem here is that ... None of your points relate to the topic, and are likely to cause topic drift as people nail you for your lack of knowledge about evolution, gratuitous assertions regarding atheists and disregard for the evidence. Please see Message 1 for what the topic of this thread involves. Enjoy.
... as you are new here, some posting tips: type [qs]quotes are easy[/qs] and it becomes:
quotes are easy or type [quote]quotes are easy[/quote] and it becomes:
quote: also check out (help) links on any formatting questions when in the reply window. For other formatting tips see Posting Tips If you use the message reply buttons (there's one at the bottom right of each message): ... your message is linked to the one you are replying to (adds clarity). You can also look at the way a post is formatted with the "peek" button next to it. Edited by RAZD, : clrty we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again Foxdog,
Any comments on the topic? You know, the issue raised in Message 1 quote: Also see
Message 2 Message 3 Message 4 for more details. This forum generally likes people to stick to the topics in each thread, and if you can't find one about a topic you want to discuss, you can start a new topic at
Proposed New Topics Thanks for your consideration. Enjoy. Edited by RAZD, : subtitle Edited by RAZD, : dbcode we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Kaichos Man, still at the quote mining and misrepresenting eh?
At any given moment you may be looking at a range of morphologies that may all be the same species. They may not be, of course. The point is you can never know. One can never be 100% certain about anything. One can, however be certain enough for practical applications, and even if one is right 90% of the time that would be more than sufficient in my opinion. You need to show that there is less than 90% confidence in the foraminifera classifications, and to date you have been totally incapable of doing that. Again what are we trying to be so certain about? The absolutely correct classification of every single variety and subspecies population all the way back to the first common ancestor population of foraminifera? Hardly a realistic goal, nor do Parker and Arnold claim any such impossible result. Certain enough that the lineages of descent are adequately described to sort most organisms into their respective places? The evidence is that this is, in fact, where we are, with some disagreements about some groups at the outlying tips of classification - whether they are species or subspecies. Certainly there has been no major revision of the taxonomy of foraminifera above the level of family and very very little at the level of genera. You have not posted a single reference that speaks to reclassification of any major level above genera. Your failure to find any such evidence shows again that we can easily have over 90% confidence in the current classifications. You're still fighting the reality here.
It seems that the "major difficulty" is sorting out subspecies versus species
No it isn't. It is:
morphological sequences that mimic evolutionary change.
"Morphological sequences" being precisely what was presented by Parker and Arnold. Curiously, your opinion has little ability to alter reality. The full quote is (italics mine for emphasis):
... clinal morphological changes due to coadaptation to similar environmental gradients can produce morphological sequences that mimic evolutionary change. In no way does this imply that this happens across the board to such a level of pervasiveness that all the data is rendered invalid. Interestingly, that would be the kind of evidence that you would need to make your claims, and the evidence still just is not there. The problem you have vis-a-vis Parker and Arnold is that these clinial morphological coadaptations within environmental gradients AND your possible ecophenotypic variations within a species in environmental gradients occur at the same time as the core species as it evolves. That core species remains in the evidence along with the varieties, and this core species allows Parker and Arnold to map their tree of relation without being sidetracked by these varieties into erroneous conclusions about these species. These variations are horizontal to the vertical geological time sequence of evolutionary change over millions of years. For Parker and Arnold to be confused by the ecoclinial variations, the core species would have to be missing from the fossil record, and sadly, for you, they aren't. Thus sorting the foraminifera out by morphology alone, as was done by Parker and Arnold, would result in branches from the main trunk. The main picture of the trunks would not be altered by what are minor variations on the theme of the core species in the overall picture. The result is an analysis with at least 90% confidence of correctly mapping the evolutionary changes over time. Now let's look at your latest misrepresentation:
Let's take a look at the way the University of South Florida views the problem:
quote:reference: http://gsa.confex.com/...5AM/finalprogram/abstract_97460.htm Sounds pretty bad at first glance, but is that really what he is saying? The full quote is:
quote: So the reason that one needs to supplement the standard reference to have a classification above the level of genera is because the standard reference (now 23 years out of date, and preceding Parker and Arnold) only classified forams to the level of genera. You forgot to mention that part, and that means your quote is a false representation, a falsehood, a quotemine.
quote: The first being 18 years out of date and the second being 11 years out of date ... ... while we have seen genetic studies since McCloskey's 2005 article that have shown that many apparent subspecies variations are in fact cryptic genetic species, where even minor morphological differences can represent real species differentiation. Certainly these new studies would improve the overall classification of foraminifera if combined on an These later studies still present a major problem for your assertions, as they have not uncovered a single case of misidentification of a subspecies that should belong to a different family. Fascinatingly, that would be the kind of evidence that you would need to make your claims, and the evidence still just is not there. Instead these studies confirm the morphological sorting down to that level of species detail, with the possibility of extending it to cover the cryptic species within current morphospecies.
quote: So once again we see that it is at the species level that the accuracy becomes a problem
quote: In other words, accurate information is available, it is just a matter of sorting it out in the quagmire of numerous different sources into a unified updated and cohesive whole. End result: over 90% validated confidence in classification down to the level of genera and slightly less confidence in classification down to species with a bit more uncertainty down to the level of subspecies variants. Hardly turning the world of foraminifera classification on it's ear. Enjoy. ps - for the record ...
Message 90: Oh, and a word to the wise. If you have come to this forum hoping to win an argument, forget it. I have never seen anyone, Creationist or evolutionist, concede defeat. Not once. When the dust settles on any given subject, both sides believe they have won. Both sides are gobsmacked that the other side can't see how badly they have lost. I've seen stubborn people who can't admit to being wrong, true, but I've also seen a number of people admit to being wrong, both creationist and evolutionist. I've seen a number of creationists being converted to evolutionists by arguments at this forum (cavediver and Rahvin are two that will tell you that they have changed as a result of this forum convincing them that creationism was wrong), but I have not seen a single evolutionist change to creationist. Perhaps you are projecting your inability to admit to being wrong onto everyone else? Edited by RAZD, : added ps we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Kaichos Man,
90%. That's not bad. 90% of 90%. 81%. Hmm. Getting a bit dodgey 90% of 81%. 73%. Oh dear. And this is the "unbroken evolutionary progression"? More than a quarter of it may be wrong? Of course you are assuming that it is 90% at each level. Remember that we are dealing with a phylum: Domain: EukaryotaKingdom: Rhizaria Superphylum: Retaria Phylum: Foraminifera This is how the taxonomy stacks up:* Kingdom If genus classifications have 90% confidence then what is the confidence levels for family, 100%? order , 111%?, and class, 123??? Rather obviously your assumed factor cannot be correct, or your application of a factor at each level is bogus. If we assume, for the sake of argument, 100% confidence for the phylum classification then the factor for each step would have to be (0.90)^1/4 or 97.4% confidence per level of classification to end up at 90% for genus: phylum 100%class 97.4% order 94.9% family 92.4% genus 90.0% Continuing on we now get: species 87.7%variety 85.4% ... better than 5 out of 6. More than a quarter of it may be wrong? Note that this is your "major difficulty" in classification in Message 55, your "[i]taxonomic quagmire[/t]" in Message 89, and it appears to be correct 85% of the time?
And this is the "unbroken evolutionary progression"? Yes, because you have absolutely failed to show that the progression they developed is completely wrong. You can't even show that it is occasionally wrong. The occasional messed up species classification does not affect the progression in transitional forms. Certainly you have not challenged the idea that transitional fossils exist and that they show transitions from ancestral populations to descendant populations. Even if Parker and Arnold were correct only half the time there would still be a mountain of evidence of transitions and a plenitude of transitional fossils. You are down to playing with fictional numbers and even there you can't show that no transitional fossils exist. Time to review where this thread started:
Message 4: In the post that began this thread Kaichos Man claimed (Message 19):
... this is prcisely what we should see in the fossil record in abundance. Darwin was certain that future fossil finds would support his theory. They didn't. So now neo-Darwinists harp on about fossils being "extremely rare", and "difficult to form". What a load of parrot droppings. Take a look around. Rivers, lakes, seas and oceans everywhere. Daily tides. Frequent droughts and floods. Fossils are being formed by the ton as we speak, all over the world. And this process has been going on for (supposedly) millions of years. If the theory of evolution was true, we would be up to our necks in transitional fossils, each tiny darwinian step lovingly catalogued in the strata. We can ascertain the veracity of this arguement from incredulity by comparing the fossil record for the Foraminifera, Pelycodus and the Coelacanth. ... Conclusions What is clear, from comparing these three cases, is:
Simply put, fossils do not need to exist to fill in gaps in the fossil record for intermediate forms to have existed. And we are still at those same conclusions, completely unaffected by any of your various arguments ("parrot droppings"). All you have done is waste posts. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1432 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Percy
This just shows how important this forum is, we have now become the first find:
Message 61: It's worse than that: when you google ecophenotype foraminifera, and this thread on EvC is the second hit. The first hit is Helen Tappan's 1978 paper. ecophenotype foraminifera - Google Search Conclusion: your forum is one of the leading experts on ecophenotype foraminifera ... or it just is not that big an issue. We are now the first pick. Helen is now in second place. Looks like the issue is so rare that the few posts here have been sufficient to alter the results. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024

 (1)
(1)