
|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| Thread ▼ Details |
|
Member (Idle past 1435 days)  Posts: 20714 From: the other end of the sidewalk Joined: |
|
Thread Info
|
|
|
| Author | Topic: Another IDology challenge -- complete with complaints of harsh treatments ... | |||||||||||||||||||||||||||||||||
|
dwise1 Member  Posts: 5952 Joined: Member Rating: 5.7 |
I'm not going to waste an entire hour listening to ID BS, especially if it's as brain-dead stiupid as you describe it.
I don't care how "learned" Gelernter's name claims him to be (German: lernen, lernte, gelernt), as a computer scientist he should know better than others the old dictum, GIGO ("Garbage In, Garbage Out") *. The output of your program can only be as good as the program and the data inputs: if your program is fouled up or you input crap data, then the results will be fouled up and crap. Or to a mathematician modeling something in the real world, your calculations depend on your model, so if you build a slap-dash fouled up model, then your results will reflect that. Building crap models and using them to throw a lot of big numbers at the audience is one of the oldest "creation science" tricks to confuse and deceive its audience, one which IDologists use fully -- it's easy to refute most YEC claims, but ID claims are more difficult because of all the obtuse pseudo-mathematics you have to wade through and counter (not to mention that your countering would also go over most people's heads). I should give Gelernter a listen before addressing his claims, but it's not worth sitting through an hour of that crap (I've had to sit through Hovind videos to find a specific claim and I have no desire to undergo such torture again). Do you have the timemark for when he makes his claims so that I can respond to what he's actually saying? Barring that, I have to go with your descriptions. For example, what does he mean by "to assemble a protein molecule by molecule"? Are we talking about DNA base pairs, triplets of which form codons which translate for amino acids? Or are we talking about amino acids which chain to form proteins? Are we talking about the original proteins or modern proteins, or does he not realize that there's any distinction? Also, is he talking about only one single attempt to form a (¿modern?) protein in such a single-step selection manner (see Chapter 3 of The Blind Watchmaker)? Or is he allowing for a large number of parallel attempts such that the attempt would succeed if even just one individual attempt succeeds, meaning that the overall attempt would fail only if each and every individual attempt fails. Refer to my page, MONKEY PROBABILITIES (MPROBS), where I analyzed the probabilities in MONKEY, my implementation of Dawkins' WEASEL program (again from Blind Watchmaker):
Another treatment of this is on my page, THE "RANDOM" PROTEINS ARGUMENT, in which I responded to a typical creationist probability claim about the chances of a modern protein just falling together randomly -- my impression is that this is what Gelernter is also trying to argue. Several problems with that:
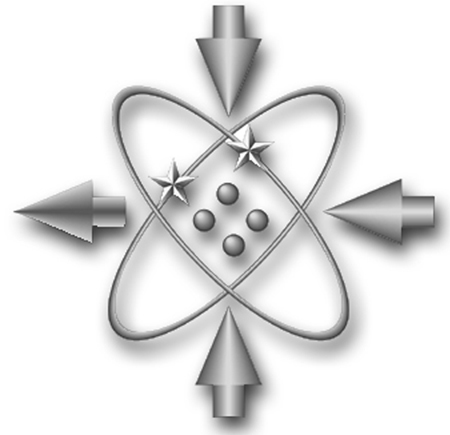
Did I misunderstand Gelernter's arguments? If so, then please give us the timemarks so I can get the straight skinny. {FOOTNOTE *: We DSes (US Navy Data Systems Technicians, disestablished in the 1998) saw GIGO, "Garbage In, Garbage Out", as being depicted in our rating symbol:
The helium atom is used in the rating symbols for all electronics ratings (not technically correct, but far easier to embroider than a copper, silicon, or germanium atom). The three arrows pointing in represent inputs and the arrow pointing out represents output. This image is of a metal pin. When embroidered on a rate badge, the input arrows are solid and the output is just an outline (empty). We would interpret that to mean that the input arrows were the unprocessed "garbage in" and the output arrow was the processed "garbage out." TRIVIA:The rating symbol for Electrician's mate is a globe of the earth. Here is why that is. The creation of rating symbols happened around 1921. The Navy team went to each rating community and asked about equipment or tools or anything else that would symbolize what that rating did. At that time, many light bulbs were spherical and were called "globes" (as opposed to our current talk of an oniony shape, "bulb", or the German "Glhbirne", "glowing pear"). So when the team spoke with the Electrician's Mate the response was "a globe", which the team misinterpreted. By the time the mistake was discovered, it was too late to correct it.}
|
|||||||||||||||||||||||||||||||||
|
dwise1 Member Posts: 5952 Joined: Member Rating: 5.7 |
So then show us! Point us to your sources. Quote from them.
I am not going to sit through an hour-long video filled with BS. I explicitly asked for a timemark so that I could hear Gelernter's argument itself. If you have something to show us, then show us.
|
|||||||||||||||||||||||||||||||||
|
dwise1 Member Posts: 5952 Joined: Member Rating: 5.7 |
I know we love to say this but I don’t think evolution has a rate. I've pointed this out before, but nobody will believe me:
Evolution never stops. The same evolutionary processes are constantly at work, just with differing results. The processes that cause changes in the population in a new or changing environment are the same processes that keep a population from changing in an unchanging environment. So even when there's no change, that's still evolution at work.
For those with a background in engineering or as a technician, it basically acts like a negative-feedback control loop. The further you are from the set-point (eg, a specified voltage, the optimal phenotype for that environment) the harder it will drive you back to that set-point. When you are at the set-point, then the exact same mechanism keeps you at that set-point. If a power supply's voltage output remains constant, that does not mean that it's not performing voltage regulation.
|
|||||||||||||||||||||||||||||||||
|
dwise1 Member Posts: 5952 Joined: Member Rating: 5.7
|
When the ecology is stable, in equilibrium, then selection is to maintain that median position because it is successful. Indeed. Selection is still selection and selection still happens. All it takes to be able to see that is some basic knowledge of evolution and thinking through how it works. The backlash I would get would mainly be from creationists who are so wrapped up in definitions and, since the word "change" appears in their definition of evolution, they think that if there's no change then evolution isn't still happening.
|
|||||||||||||||||||||||||||||||||
|
dwise1 Member Posts: 5952 Joined: Member Rating: 5.7
|
I like the analogy. I'm going to steal it. Thanx. You're welcome to it. I'll even give you some of the background to its development in case that gives you more to work with. I read an article from Nature or a similar science journal from around 1980 which reported on a paleontology conference centering around punctuated equilibria and similar topics. One pattern presented would be of a large population ranging over a broad environment such that it didn't change much but it lasted a long time because it had enough diversity to weather through changes in their environment; compared to that are smaller populations changing a lot to become highly specialized to their particular niches, but short-lived because they could not survive changes in their environment. A graphic that formed the imagery in my mind was one which showed that the "sudden" change in geological time was still gradual in generational. The portion of the graphic showing generational time was a series of bell curves each representing a generation and showing the center of the curve (representing the optimal organism) moving as the environment changed. That would translate into our use of the term "set-point" here and of your point that that set-point does move so the population needs to track that movement. Another contribution was made by a PBS popularization of evolution from the early to mid-80's hosted by Christopher Reeve. Towards the end was my first exposure to Evolutionstechnik, using evolutionary processes in engineering (eg, genetic algorithms, it was a few years later that I first heard of GAs). The mind-bender for me was when he presented a model of a 3-D "evolutionary landscape" in which the environmental optimum was the top of a hill (a local optimum) and he described evolutionary change as being faster when the population is farther away from that optimal point and would slow down as it got closer. That idea took me by surprise, so I worked through some Gedankenexperimenten (I'm kind of good at visualizing things). This is what I ended up with (which would work better with good visuals, so my apologies):
In my mind, I saw that population work its way to that optimal point and then center around it. And over subsequent generations as the population's curve might try to spread out, selection would eliminate the more extremely different individuals and thus keep the population centered at that point. A further application would be to start with an optimally adapted population and then start moving the optimal point and observe the population's response to that. Again, we should observe the population shifting its own center to track its optimal point. I think that you could set up an experiment involving two optimal points, one that the population starts off tracking and the other a near-by one. I would visualize part of the parent population splitting off and starting to track the new optimal point. I've also thought of adapting this visual model to a simulation program for study. Let us know what you're able to do with this, if anything. Share and enjoy!
|
|||||||||||||||||||||||||||||||||
|
dwise1 Member Posts: 5952 Joined: Member Rating: 5.7 |
Faith writes:
Again this is not how evolution works. It doesn't blend characteristics. It selects those that are more fit for survival or more attractive for reproduction. That's what selection does. It will eventually blend tog3ether whatever proportions of traits are in the new set of individuals, their new collection of gene/allele frequencies, and eliminate others from the population. It appears to me that Faith is making the same mistake as Charles Darwin when he couldn't figure out how new traits could establish themselves in a population. He pictured heredity as being like mixing paint, but that would inevitably result in new traits disappearing as all traits just blended together. But of course, that's not how it works as scientists came to realize through Mendelian genetics (ironically, Darwin had a copy of Mendel's monograph in his library, but apparently never got around to reading it). We now mainly only know about Darwinian evolution (the natural selection acting upon variation part), but we forget about Darwin's pangenetic theory which tried (and failed) to explain heredity. We have reams of creationist quotes of "scientists declaring Darwinan evolution to be false" which are taken primarily from the first part of the 20th century. During that time, research into genetics and mutations did indeed disprove Darwin's pangenetic ideas (which were demonstrably wrong), so you can find many statements of "Mendel proves Darwin wrong!", but that applied only to pangenesis and had nothing to do with the natural selection part. By the 1940's, it was discovered that Mendel actually provided answers to the questions that Darwin couldn't answer himself. What resulted was the Modern Synthesis (AKA "Grand Synthesis"), the combining of genetics with Darwinian evolution which produced neo-Darwinism. So all that Faith has succeeded in doing is in reproducing Darwin's biggest mistake.
|
|||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024

 (1)
(1)