|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| EvC Forum active members: 65 (9164 total) |
|
| |
| ChatGPT | |
| Total: 916,914 Year: 4,171/9,624 Month: 1,042/974 Week: 1/368 Day: 1/11 Hour: 0/0 |
| Thread ▼ Details |
|
Suspended Member (Idle past 1475 days)  Posts: 35298 From: Nevada, USA Joined: |
|
Thread Info
|
|
|
| Author | Topic: Evolution Requires Reduction in Genetic Diversity | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days)  Posts: 20714 From: the other end of the sidewalk Joined: |
... But the other one makes a weird detour all the way around the aorta in the thorax only then returning to its final destiny in the neck again. In giraffes this causes an astonishing detour of several meters. Yep, that is on my list for another post on the Silly Design thread.
It must have pleased Him. A real artist He is. Certainly more like an artist than an engineer ... Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
Faith: But when I mentioned the failure to interbreed occurring at the extremes of a series of population splits, that was a direct challenge to the idea that mutations would be the cause, because there is no reason why they would affect only the extremes where it's the normal recombinations that would have the major effect. trochiloides is the ancestral species. Now imagine there are 10 mutations between trochiloides and ludlowi - not enough to cause infertility. There is also 10 mutations between trochiloides and obscuratus so they are able to hybridize freely as well. Also 10 mutations between ludlowi and viridanus and between obscuratus and plumbeitarsus, all able to freely hybridize. Thanks, I just do not see how dividing alleles into different populations can result in genetic incompatibility. The alleles are not the genes, but just a part of them. In a population with four alleles: A, B, C and D you can have theses combinations:
If we split the population into those with A and B and those with C and D then each subpopulation can have the following combinations:
So then if we reintroduce the two populations to each other ...
None of those offspring are any different mix than occurred in the parent population, none of those parent varieties did not occur in the parent population. Without any change to the gene how could there be genetic incompatibility? Without any change to any of the alleles, how could there be genetic incompatibility? Even with change to create a new allele, how could there be genetic incompatibility? If we assume a new allele E in population 2 then we get these possible offspring:
So the recombined population would then have:
There are only two new combinations between pop1 and pop2: AE and BE, so -- unless the of the gene is changed, how could there be genetic incompatibility when the populations recombine? As I see it there is the basic function of the gene, and there are variations that don't affect function but do affect the phenotype (ie alleles). As I see it, to get genetic incompatibility you need to affect the basic function of the gene. Or do I have this wrong? Enjoy Edited by RAZD, : resorted table for clarityby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Is anybody going to dispute that a small population is inevitably going to develop genetic drift which DOES change allele frequencies? ... Of course not --- ANY population is inevitably going to develop changes in allele frequency due to genetic drift, it is just more noticeable in small populations. We just know that it is one of many mechanisms that affect evolution of species within habitats.
And is anybody going to dispute that inbreeding can multiply undesirable traits simply by pairing recessive alleles (normal genetics)? Again, of course we won't --- ANY population is inevitably going to pair recessive alleles through normal mating, it is just more noticeable in small populations. As herebedragons says: founder effect. Any small population without significant selection pressure will see more of the homogenic recessive offspring survive and interbreed than a larger population. We just know that it is one of many mechanisms that affect evolution of species within habitats.
... The newly isolated subpopulation won't look any different from the population it split from until these new combinations are worked through for some generations, though if the population is quite small some dramatic individual changes could show up sooner, maybe even in the first generation. And that's still my main interest. That doesn't explain genetic incompatibility.
I did think that the new combinations could lead to a genetic mismatch preventing interbreeding in some circumstances, such as by pairing recessive alleles for instance, but it isn't important to my argument. What new combinations? This is the combinations you can have before the split:
And that is also the combinations available when reunited -- unless one allele is mutated (D → E in my example). Nor do I see how a new mutated allele can affect genetic compatibility. The possible combinations post mutation are:
There are only two new combinations between pop1 and pop2: AE and BE, so -- unless the of the gene is changed, how could there be genetic incompatibility when the populations recombine? What I am trying to do here Faith is draw a line between changes to alleles and changes to gene function. It seems to me that only changes to gene function can cause genetic incompatibility.
I did think that the new combinations could lead to a genetic mismatch preventing interbreeding in some circumstances, such as by pairing recessive alleles for instance, but it isn't important to my argument. What I don't see is how shuffling existing alleles changes gene function enough to cause genetic incompatibility. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
... In the case of the greenish warblers, IIRC, where the two ends of the ring meet the two species have different songs and so do not recognize each other as potential mates. Could they produce viable hybrids if forced to mate? Maybe, but does it matter? In the latest study they did find some (rare) hybrids between the two northern varieties, so we know that genetic incompatibility is not involved, just shuffling of alleles and some changes to alleles, not to gene function. This of course also means that according to the biological definition of species that trochiloides is in fact all one species.
2) Prezygotic isolation - ie. different pollinators, different breeding patterns/timing, different mating habits (bird songs/displays) It would seem that this is what is affecting trochiloides as it appears they migrate south to similar locations but spread out in these locations to breed.
1) Ecological or geographical isolation Is actually just one way that Prezygotic isolation occurs, so I would combine (1) and (2).
Genetic incompatibility can come from a number of things. It could be from incompatible genes or from failure of the chromosomes to form viable pairings during recombination or from changes in regulatory networks. (I suspect that the latter to be the most common). This would occur via mutations that change the bend sequence\shape of the DNA strand, which would also change the function (protein production) of the gene yes? If that pairing incompatibility were due to a recessive gene, then would not that very same pairing incompatibility prevent that gene to participate in heterozygous mating?
So in theory, one mutation could result in prezygotic or postzygotic incompatibility but it is highly unlikely. It more probable that it is due to a series of incompatibilities that accumulate. The only cases of single mutation speciation that I am aware of involve polyploidy.
The population genetics of speciation: The evolution of hybrid incompatibilities Also check out this paper Hybrid incompatibility and speiation We can talk about them later. Indeed. I am reading the first, and note this comment:
quote: (figure 1 is on p 1806 and shows the divergence of two theoretical populations with genetic mutations, ie - a → A, b → B, etc) This basically is the potential for genetic incompatibility and requires at least two exclusive mutations, and the first of the untested compatibilities is AB. These untested compatibilities do not occur with all existing alleles in a population. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
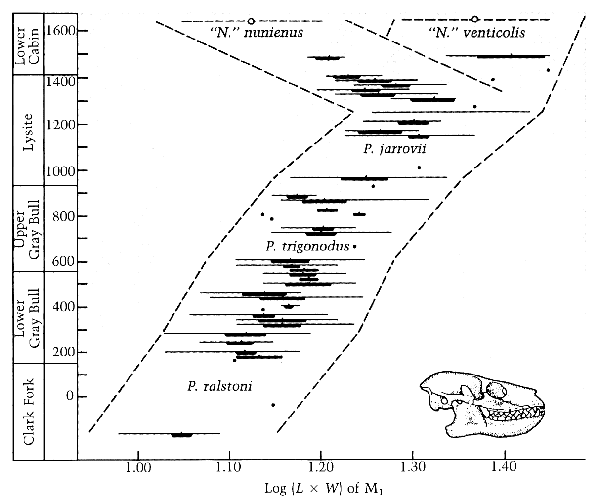
The original chart gives what sound like the names of locations on the left margin. Were these fossils found scattered in many locations, or in roughly the same geographical area or even in a column one layer above another? Or do the names designate the layers themselves. ... My understanding from reading other articles is that it is the same general area, but not all in one column, and they are grouped by the geological layers they are found in, the names are for groups of layers, just as is done in the Grand Canyon. There are other nearby areas that have also been excavated without finding this lineage (I can provide more if you are interested). btw the thick part of the lines represents two standard deviations within each population ... so 95% of the fossils are grouped in the thick sections at each layer. The thin part of each line represents the full range of fossils from largest to smallest at each interval\layer within the remaining 5%. Here is the transcribed chart again for reference:
... And does it matter how many fossils were found in each layer? Yes, because the more fossils found in a layer the better representation you have for the variations within the population.
So this is all about the size of the creature. Yes, they are so similar that you can mix fossils from neighboring layers that are in the same size range and then not be able to sort them back out with confidence.
This layer is usually dated to such and such MYA. The many collections of these fossils found in various layers appear to differ in average size from one layer to the next. The measured sizes are such and such." And the dates each was found could be handy, as well as who found them etc. ... Curiously what you have is the data that show just that, albeit with relative age from the layers, not specific dates from any other measurements.
... Instead of simple description we get theory. Why is that? What theory Faith?
No problem with superposition of course, tells the order they were buried in ... and thus the relative ages of the fossils.
... which might have something to do with their size though it might not; but geological time is always problematic. Which you know is not true due to the strong degree of overlap in sizes, and because of the time between deposits: why would there be 80% overlap in sizes from one layer to the next if there were some magical sorting mechanism? Why would animals the same size be sorted into different layers? How would animals the same size be sorted into different layers?
Again this is of course evolutionist theory. ... No Faith, it is looking at -- reading --what the evidence tells us: similar animals in discrete time differentiated layers, layers that have to be from different times laid one on top of the other. You acknowledge that superposition means upper layers came after lower layers
... If they were simply buried all at once in a one-time catastrophe the only continuity would be that they are clearly the same species that got sorted into different burial locations for one reason or another. ... Again, why would there be 80% overlap in sizes from one layer to the next if there were some magical sorting mechanism? Why would animals the same size be sorted into different layers? How would animals the same size be sorted into different layers?
... Did anyone differentiate between adult and young individuals or male and female? They were differentiated by age (teeth).
It could. Since four subspecies/races are noted the sizes could reflect the range of genetic diversity for size in each population, which I think is what you have in mind. But again I'd like to know if the size could be related to age or sex and if that was taken into account. Those labels are arbitrary human divisions for the purpose of discussion, based on the layer time intervals. Some other documents break the lineage into more divisions (ie - P. frugivorous just below N.nunienus).
First, of course I dispute the whole evolutionist interpretation that says the populations genetically diverged from each other over time as represented by the layers, which I guess would make P. ralstoni the ancestral population. ... It is what the data shows. We know they are related because you could mix the same sizes from adjacent layers and not be able to sort them with confidence, and we know that the upper layer was deposited after the layer below it.
... My interpretation is that they are certainly all related ... Agreed.
... and may be different subpopulations ... Mundanely true in that the same animal cannot become two fossils.
... but that they would have been contemporaneous with each other, ... Which is at odds with the evidence. Thus your interpretation fails to explain the evidence.
... probably inhabiting different locations. ... Also at odds with the evidence that they came from different excavations in the same general area, and that adjacent areas have fossils of similar sized near relative Copelemur fossils but not Pelycodus fossils. So I will agree that Copelemur was a contemporaneous population inhabiting different nearby locations, morphologically similar but distinct enough to separate.
... If some are subpopulations of others there is no reason to think they lived in different time periods anyway, or that the higher in the strata are necessarily the offspring of the lower: it could be the other way around. Riiiiiiiiiiiiight, the offspring are somehow buried below the parents???
But for the sake of argument, accepting the evolutionist interpretation that the higher descended from the lower over time, and that size was the main variable from population to population, ... That is what the evidence shows.
... while there should always be a trend to loss of genetic diversity as a new subpopulation develops its new gene frequencies, there's no way to know, especially from fossils, whether the alleles for the other trait completely disappeared from the new population. ... All we can know is what the data shows, but after several layers of consistent shift to larger sizes and no outliers (the thin lines show the full extent from largest to smallest) we can have confidence that certain size alleles are no longer expressed.
... If they still exist in the collective genome, ... Then they would be expressed. Remember that the thick parts of the lines encompass 95% of the fossils found, that the thin lines extend to the largest and smallest fossils in the remaining 5% so the probability of single individuals being significantly outside these limits is very very small. When you consider this small probability recurring at each level the possibility becomes vanishingly small.
... further population splits could select those alleles, ... Precisely as seen at the top of the chart where there is the initial division. Population splits can only select alleles within that population -- selection does not create alleles.
... and two separate populations branching off could select alleles for each end of the spectrum, which would be reflected in the size differences between the two groups at the top. ... That is what you see at the time of the split, but then each population continues to diverge by developing sizes that are NOT in the parent\split population. The large group continues to grow to sizes not seen in previous populations with magical new size alleles, the small group revisits the sizes of the bottom population, but not seen in intermediate populations. If we consider this a test of your hypothesis that alleles are permanently lost then the evidence shows your hypothesis to be false. On the other hand if you now hypothesize that alleles from the parent population not seen expressed in the later populations are hidden but still exist in the "collective genome", then your hypothesis become useless because nothing is truly lost it just gets magically hidden until needed.
... Or those two populations could reflect incompletely inbred genes, since the first stage of the expression of new gene frequencies is likely to bring out a motley collection of traits, possibly including the extremes, which would work through the population through many recombinations over time until a general phenotype emerges. Irrelevant to your problem. There would be a founder effect, but each population is large enough that inbreeding would not be a problem. Once they have divided selection would drive them further apart to lessen competition. The problem remains that the left hand small group branch recovers alleles that should have been lost if your hypothesis were correct.
So there would have been a reduction in genetic diversity if these really were subpopulations formed over time, ... It isn't necessary, but it does happen. Here we see about a 50/50 split in size but a large variation remaining in each population -- as large a variation as seen in lower populations.
... but complete loss of alleles for different traits doesn't necessarily always occur. ... In other words you have made up a "get out of jail" card that you can play every time your hypothesis is shown to be falsified by empirical evidence.
... but complete loss of alleles for different traits doesn't necessarily always occur. It's a TREND I'm trying to highlight, that only becomes expressed in complete genetic depletion at the end of a series of subpopuloations. It is a trend that has not been observed. Ever. What we see is occasional temporary depression of diversity at the time of population division, followed by increase in diversity within each daughter population as new alleles are added. By Mutation. Even when there is a very small founding population with very few alleles. The two populations at the top each have as much diversity as the populations at the bottom.
I don't think that is what happened though because of course I don't accept the whole evolutionist interpretation that these fossils represent different populations that formed at separate times from the one lower in the strata. Of course, because you don't accept actual evidence in place of imagination, especially when it invalidates your imagination. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Oh my, do we really need to explain why or how upper layers come after lower ones? We cannot even refer to obviousness here. Weird.And absurd. No, Faith does understand the law of superposition, but she has a tendency to discard inconvenient facts when the counter her thinking, as you may have noticed. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
Reproductive isolation is any barrier that prevents or reduces gene flow between two populations. ...
Speciation cannot be thought of as an event, but as a process. ...
Genetic incompatibility is when there is a genetic basis for the inability of the hybrid offspring to contribute to the recombining of the parent populations. ... Now, I will admit that this is my own definition so I would be happy to debate it but I'll give some justification here for this definition. These work for me. Isolation initiates the process and the fixation of genetic incompatibility (post-zygotic separation) completes the process. Isolation may be over many many generations and over widely separate intermediate locations, with populations accumulating mutations, but genetic incompatibility isn't a foregone conclusion, especially if there is no stress/selection to change gene functions.
I think it is pretty obvious that if the hybrids are not viable, (ie. they don't develop into adults) that they will not themselves breed and produce offspring and so cannot contribute to recombining the parent populations. It should also be obvious that if the hybrids spread through the population and completely homogenize it, they are NOT genetically incompatible. However, in-between these two extremes is a whole gradient of possibilities. Indeed. With the Asian Greenish Warbler Phylloscopus trochiloides we see separation and variation\modification resulting in a low occurrence of hybrids at the ring end overlap area, due to behavior\song\mating choice, but no definitive evidence of genetic incompatibility ... ... however there could be reduced viability if some alleles are lethal but others are viable, and that could be a factor in the low recording of hybrids). From your H. Allen Orr paper The Population Genetics of Speciation: The Evolution of Hybrid Incompatibilities:
If the left population had some (a) mixed with some (A) alleles, and the right population had some (b) mixed with some (B) alleles, then you could get:
ab, aB, Ab and AB Of these only AB is not tested and could be lethal\sterile, and if so, then reproduction would be depressed compared to normal fertility, with the degree depending on the relative proportions of a:A and b:B in the breeding populations. And of course there could also be interaction with other genes that could affect viability\fertility of the hybrid. And some could be sex-linked so male (A) could mate successfully with female (B) but male (B) would be lethal\sterile when mated with female (A). It comes down to the fixation of the mutated genes in the populations and their effect on viability\fertility when the two populations meet.
Here is an example. Let's say we have a population of plants that is adapted to higher elevations and one that is adapted to lower elevations. Where these two population meet there is a hybrid zone. However, the hybrids are not well suited to the higher elevations; but neither are they well suited for the lower elevations. So the hybrids remain confined to this narrow zone and the two populations retain their distinct character, thus gene flow is restricted to a small section of the two populations. This would be considered partial incompatibility, but it would be enough to allow each population to remain differentiated. We could even measure the amount of partial incompatibility by creating a cross and transplanting the hybrids to both habitats and measuring fitness. A reduction of fitness by say 90% in each habitat would indicate a 90% incompatibility. Exactly.
No, I wouldn't combine them. They are two different ways of restricting gene flow. Prezygotic isolation could occur without geological isolation and geological isolation could occur without producing prezygotic barriers. Well I think they are just different forms of isolation, and I would list it
... we can agree to disagree, but the way I see it is that there are several ways that subpopulations become isolated, and that it is the isolation that is critical for further genetic divergence. An example of (3) would be your Monkeyflowers, where different pollinators enable the isolation. An example of (4) would be the Asian Greenish Warbler in the overlap zone
Here is a study on Monkeyflower where a substitution at one locus (YUP) caused a dramatic shift in pollinator preference and sympatric populations are now 99% isolated - because of this one mutation resulting in prezygotic isolation. They will hybridize in the lab, which is how they made their NILs (near isogenic lines - which is single gene from one species introgressed into the background of the other). But they are reproductively isolated in nature. With the Monkeyflower we see a genetic basis for isolation by causing a shift in how they interact with the ecology (pollinators) while not being genetically incompatible.
The factors that prevent most of the two northern varieties from interbreeding are probably genetic (although bird songs can be learned from the parents, so if that were the only difference then it may not be genetically based). So perhaps it could be partial genetic incompatibility but unless we know something about the hybrids, more likely simply prezygotic isolation. With the Greenish Warbler we see a genetic basis for isolation by causing a shift in mating preferences (song and wing pattern), while not being genetically incompatible. So the genetic isolation can be initiated with a genetic change without necessarily resulting in genetic incompatibility.
[qs]Genetic incompatibility can be very simple as in the case of Mimulus quote: (For clarity these are different monkeyflowers than the other paper above.) So we have m1f2 sterility but m2f1 (reduced) fertility, interesting.
The following chart is adapted from a lecture on DMI (Dobzhansky—Muller Incompatibilities) in my Evolutionary Biology course.
It shows how one substitution causes no incompatibility but the number quickly rises as the number of substitutions increase. hmmm ... the "V" diagram above is faster growing
(potential) incompatibilities(n) = Σ(subtitutions, s=1→n){s-1} and it doesn't matter which branch the substitutions occur on.
So in the case of this monkey flower (Mimulus) there are substitutions at 2 loci, hms1 and hms2 resulting in 1 incompatibility which causes almost complete male sterility in the hybrids. And the possibility that one additional substitution enables genetic incompatibility increases dramatically as the number of substitutions increases. What we don't know is how many substitutions are involved in the initial split of the two species populations. For instance if we are at (e) on one branch and have (E) on the other,
then even though the other substitutions did not affect viability\fertility the latest one has 4 opportunities to do so. Thanks.by our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
In extreme cases, the founder effect is thought to lead to the speciation and subsequent evolution of new species. YES! Seems to be saying that the new allele combinations ALONE are sufficient to lead to speciation, contrary to some pretty aggressive assertions on this subject in this thread. No. The founder effect is the platform on which following populations are build. In order for speciation to occur there must be a genetic difference that inhibits interbreeding when populations reconnect. That difference is shown to be due to different fixed mutations. A small population is more likely to fix more new mutations to be shared as they need every reproductive individual to grow. Here's that "V" diagram discussed above:
We can imagine that the left branch is a small founding population and the right branch is the parent population. Even with these mutations becoming fixed in each population, genetic incompatibility is not guaranteed, but without them you don't have any possibility of genetic incompatibiliity, as then all possible combinations are tested. If some combinations of alleles is lethal then it will continue to be lethal. If some combination of alleles is infertile then it will continue to be infertile. Any other combinations get distributed within the population. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
I was entering this thread three weeks ago. Since then I am still awaiting answers by Faith. I think EVC is no place for me.I am looking for debate and not the usual and habitual dodging and ignoring. Curiously I've been on this forum for a little over 10 (yikes) years, and Faith has been here longer ... nothing really changes. What you come to realize (if you don't leave in frustration) is that Faith provides a sounding board that allows us to discuss specific points in detail, whether is is the geology of the Grand Canyon or the genetic basis of evolution. And you come to realize that the point of the debate is not to convince Faith, but to lay out a cogent argument bolstered by empirical evidence so that people lurking can follow -- they are your true audience. Personally I enjoy learning new things from others from their responses. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
The quote on Wikipedia seemed to say that speciation can occur from the normal combinations of alleles present from founder effect so you appear to be arguing with that quote. Difficult to know which wiki article you refer to without links, so I am going to assume from other conversations (correct me if I'm wrong) that you mean: Founder effect - Wikipedia
quote: ... as I know that you will be drawn like a moth to flame by the phrase "the loss of genetic variation" ... The problem is that initially those (all red) or (all blue) populations may appear different, they are not yet different species -- those combinations could and did appear in the parent population, and thus there is no way that they were genetically incompatible with others in the parent population --- yet.
The quote on Wikipedia seemed to say that speciation can occur from the normal combinations of alleles present from founder effect so you appear to be arguing with that quote. For one, Wikipedia is not a source of absolute knowledge, but for two what it says is not exactly what you have been arguing:
quote: It doesn't cause speciation, in extreme cases it can lead to speciation. How? What I said in Message 686 was (emphasis added):
quote: Besides, you are assuming mutations without showing that there are any mutations there. ... Yes, because the preponderance of evidence shows that mutations occur in every generation, that rarely do offspring have the exact same genetic elements of their parents, because the sun rises every day whether you see it or not. Many do not show up in phenotypes, but are platforms for later mutations that do, and mildly deleterious mutations can still be propagated if the phenotype as a whole is able to survive and breed.
It's often said that mutations simply do not appear frequently enough to have any kind of beneficial effect so how can you assume they're occurring here? You seem to be confusing the appearance of beneficial mutations with the occurrence of all mutations. Most appear to be neutral (intially) or have negligible effect, and thus many are propagated because the whole phenotype is selected. Beneficial\deleterious are also often relative to the ecology (while viable\lethal are related to the individual), and when the ecology changes their relative importance to the survival of the individuals can change -- deleterious can become beneficial, beneficial can become deleterious (pocket mice and lava beds).
Why do they have to be fixed "mutations" anyway? The idea seems to be that fixed RARE alleles is the point, why do they have to be mutations? Let's start with what "fixed" means ... from fixation (wiki) quote: So you can't have "fixed RARE alleles" as a fixed allele is the only allele.
Why do they have to be ... "mutations" anyway? ... Because that is how you get something different.
Why do they have to be fixed ... Because (a) that is what happens over time, and (b) genetic incompatibility arises between fixed alleles in a breeding population. If you have 'a' (original) and 'A' (derived) alleles in a population and 'b' (original) and 'B' (derived) alleles (at a different loci) then the mating possibilities are:
ab, aB, Ab and AB If a mutation arises and is not compatible within the breeding population then it perishes from the population. So of these possible combinations only AB is not necessarily viable while the others necessarily are. If this were within a single breeding population you would have a potential for depressed reproductive\survival results, but not genetic incompatibility, and this would tend to eliminate the new mutations over time. But with population isolation the new mutations that spread in each population are backwardly compatible with the (other loci) alleles of their parent populations. Getting back to the "V" diagram again:
When each branch evolves independently with different new alleles becoming fixed within their populations they are each backwardly compatible with the (other loci) alleles of their parent populations, but not necessarily compatible with different new fixed alleles in the other population, as that is not tested within either subpopulation. That opens the door for genetic incompatibility to occur ... that is what makes incompatibility possible. Enjoy Edited by Admin, : Fix dBCodes.by our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
Just a small point Faith,
... Evolution, meaning macroevolution, meaning evolution as understood by the Theory of Evolution, really is impossible. Macroevolution in science means (and always has meant) speciation, and the formation of nested hierarchies. That this is impossible would astound several people who have actually observed speciation and the formation of nested hierarchies.
(1) The process of evolution involves changes in the composition of hereditary traits, and changes to the frequency of their distributions within breeding populations from generation to generation, in response to ecological challenges and opportunities for growth, development, survival and reproductive success in changing or different habitats. This is sometimes called microevolution, however this is the process through which all species evolve and all evolution occurs at the breeding population level. If we look at the continued effects of evolution over many generations, the accumulation of changes from generation to generation may become sufficient for individuals to develop combinations of traits that are observably different from the ancestral parent population.
(2) The process of lineal change within species is sometimes called phyletic speciation, or anagenesis. We saw this pattern with Pelycodus where different species were defined as we went from P.ralstoni to P.jarrovii
If anagenesis was all that occurred, then all life would be one species, readily sharing DNA via horizontal transfer (asexual) and interbreeding (sexual) and various combinations. This is not the case, however, because there is a second process that results in multiple species and increases the diversity of life.
(3) The process of divergent speciation, or cladogenesis, involves the division of a parent population into two or more reproductively isolated daughter populations, which then are free to (micro) evolve independently of each other. The reduction or loss of interbreeding (gene flow, sharing of mutations) between the sub-populations results in different evolutionary responses within the separated sub-populations. Each population then responds independently to their different ecological challenges and opportunities, and this leads to divergence of hereditary traits between the subpopulations, both to adapt to different ecologies and to reduce competition, as seen at the top of the Pelycodus diagram. Scientific macroevolution has occurred, it has been observed, and it is a fact that speciation and the formation of nested hierarchies does occur. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
But if speciation is not really a new species but just a subspecies that has lost the ability to interbreed ... Losing the ability to interbreed is the definition of speciation, and this is why people are complaining that you are trying to redefine the terms. Personally I don't see what the problem with new species is for creationist\creationism, as no-one claims that "kinds" are species as far as I know. We have discussed this before, and I have brought up cladistics before. At it's most simplest level, Cladistics is defined as:
quote: You can start with the "original" common ancestor from the day of creation, or from the "kinds" preserved on the ark, as a creationist, or you can start with species alive today and work your way back to see how far it goes (does it go all the way to "original" breeding populations of archaic bacteria?), as a biologist. The process is the same -- microevolution and speciation. For example: quote: This could be the "bear kind" clade for a creationist, or part of the overall tree of life for a biologist.
... and if as a matter of fact it possesses reduced genetic variability, then it is sheer illusion to call it speciation. Whether or not it possesses reduced genetic variability, speciation is defined by the loss of ability to interbreed. Complaining about a word being used in a science the way scientists defined it is sheer pointless madness -- all you will do is cause misunderstanding when you use words to mean different things than the say other people are using them. This is why Percy is asking you to clarify your position. Consider how many times you complain about people not understanding your position, and then consider that it is because of the way you have said things and the words you use.
Which is not based on any kind of evidence of relatedness between the different groups but is only an appealing mental construct imposed on the facts. Again, speciation (the loss of ability for two subpopulations to interbreed) has been observed and the formation of a clade at its most simplest level is
|
^ speciation
/ \
/ \ This clade includes the parent population and the two genetically incompatible daughter populations. But it doesn't matter whether you call them species, subspecies, subsubspecies or whatever -- it is a fact that this occurs, as it has been observed.
I suppose it would. But those things aren't impossible, they just aren't what they are interpreted to be. They are actual factual daughter populations that have actual factual evolved from a parent species and are actual factual genetically incompatible. Does that meet the scientific definition\usage of speciation? Yes: ergo speciation has occurred, no interpretation needed.
(1) The process of evolution involves changes in the composition of hereditary traits, and changes to the frequency of their distributions within breeding populations from generation to generation, in response to ecological challenges and opportunities for growth, development, survival and reproductive success in changing or different habitats. Yes, that's the typical definition I'm challenging. Again, why? This says nothing about whether genetic diversity increases, decreases or stays the same. This is the way scienced defines (micro)evolution - which you accept occurs. Whether or not it possesses reduced genetic variability afterwards, evolution is defined by the changes in the composition of hereditary traits, and changes to the frequency of their distributions. Complaining about a word being used in a science the way scientists defined it is sheer pointless madness -- all you will do is cause misunderstanding when you use words to mean different things than the say other people are using them. This is why Percy is asking you to clarify your position.
And they do, through normal microevolution as I've been laboriously describing it, ... Exactly. Microevolution is the only evolution that occurs. It is how species change over time and it is how speciation occurs when daughter populations lose the ability to interbreed when microevolution in isolation from each other results in genetic incompatibility.
... which reaches a natural end point where macroevolution should begin but can't because of genetic depletion. And this is where you run into trouble using words to mean something else than the way it is defined in science: the formation of genetically incompatible daughter populations IS macroevolution to scientists, so we know that macroevolution occurs because this has been observed. Can you tell me what you mean by "macroevolution" (presumably something different from the scientific usage\definition)?
Have you read any iof my argument at all? Yes, Faith -- reading it and disagreeing with it does not mean I haven't read or understood it, just that I don't agree. And when you say things that are contradicted by facts I will continue to point that out.by our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
I read this and can't help but think of the insect world. Genetic changes have created such a vast array of insects, many of which are (in many cases) MUCH more complicated than mammals and other vertebrates. A good example is the lightning bug. No mammal has developed bio luminescent capabilities. And then there is the issue of cryptic species -- populations that look alike but can't interbreed. The malaria carrying mosquito is an example: when they were looking for biological control methods they discovered that one kind did not carry the bug and the other did, even though they looked the same. One population bred at dawn and the other at dusk, and a slight variation in the genitalia prevented interbreeding. Species complex - Wikipedia
quote: Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
I don't know yet if I need to continue to argue against the definition of speciation, but I'm leaving it for now. So inability to interbreed just IS the definition of speciation. Look at it this way Faith: we observe that populations split into subpopulations inhabiting different habitats, they become reproductively isolated ... over time their genetic changes as they adapt to these different habitats produce alleles that cause genetic isolation when they get acquainted. We need a name for this, and the name chosen is "speciation" for convenience. It also matches the biological definition of species as being a breeding population that is reproductively isolated from other breeding populations. The name is arbitrary human identification of a process that exists whether we name it or not. Now you have been trying to say that this happens with subspecies ... a distinction without a purpose that I can see. In cladistics there is not the emphasis on genus and family and all the other taxonomic labels, partly because at the point of separation\genetic isolation you have "species" -- another label for convenience of human communication. All organisms are members of a species breeding population, whether that population has diverged into many daughter, granddaughter, etc, populations.
|
^ a
/ \
/ \
/ \
/ ^ b
c ^ / \
/ \ / \
d e f g Where a, b, and c are speciation events, and we can label their parent populations a, b and c as well. Population a gives rise to populations b and c, population c gives rise to populations d and e and population b gives rise to populations f and g. They are all species as far as cladistics is concerned.
I still have the question, What if speciation brings about new species that are always genetically less diverse or variable than the populations that preceded them? Doesn't that imply LESS ability to evolve and doesn't that imply that it doesn't really fulfill the evolutionary requirement assigned to it in its official definition:
Speciation is the evolutionary process by which new biological species arise. That is, if new biological species CAN'T arise from this point due to genetic impoverishment, what then? The offspring are always members of a species, by definition. From my side of the argument there is never a point at which mutations cannot provide new genetic diversity and replenish the well.
Sure, but if it has less genetic variability with which to evolve beyond this point, how is it "the evolutionary process by which new biological species arise?" If you have speciation -- two populations that become genetically isolated -- you have new species by definition.
You know you have a new population that can no longer interbreed with others and you are calling that macroevolution, which implies the ability to continue to evolve beyond that point. Not quite, I call the long term changes in the populations and the isolation and development of genetically incompatible daughter populations -- the formation of a clade -- macroevolution. It occurs via evolution within disparate populations. Macroevolution is looking at the larger picture of all species, while microevolution is looking at the changes in a single breeding population. As I see it there are two options for a species -- continued evolution (because nothing is static) ... or extinction. As long as you have mutations and selection mechanisms, evolution will continue to occur.
I think evolution comes to a natural stopping point with the loss of genetic diversity brought about by the very processes that produce new species. I think of this as the outer limit of the Kind beyond which no further change/variation/ evolution is possible for sheer lack of the genetic stuff needed for it. It's more of a functional definition than a category definition. If there is a natural ending point then you can never get a new "species" or the species you do get is a dead end in itself and not a platform for further evolution. Curiously what you think has little effect on reality. To move beyond the hypothetical concepts you need to show evidence that such decrease in viability occurs. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
If you put a hundred human beings on an isolated island where they have children and grandchildren for a couple hundred years, when you revisit them they will have acquired a look that is completely their own, and that will be based completely on the particular set of alleles shared among them, that excludes who knows how many alleles they left behind them in the human population at large. They have created their own race or subpopulation from limited genetic diversity. That HAS to happen when a population is started from a smallish group. That is in effect what has happened with the dispersal of Homo sapiens out of Africa and spreading in small bands to eventually cover the earth. Polynesians in particular would fit that model. North American natives meeting European explorers are like the closing of a ring species, from different groups spreading around the world. The changes in the different populations appears to be more a result of genetic drift than selection, as it would seem that eye color and shape have little to do with survival or reproduction (other than some element of sexual selection), and skin color seems to be mildly selected by latitude and UV\vitamin D effects (it doesn't prevent people of different skin color from living anywhere). Certainly Europeans and native Americans were reproductively isolated for many generations, evolving unique traits, but also demonstrating that reproductive isolation does not necessarily result in genetic isolation without some selective pressure to change.
Darwin's Galapagos tortoises got their own look simply by starting from a limited number of individuals that were isolated from the mainland population and working through the particular set of alleles they happened to possess for whatever number of generations they had been on Galapagos. This is really how evolution proceeds, it ALWAYS involves the loss of alleles as a specific set of allleles becomes the basis for a new population. And even on different islands: each island has a different species of tortoise, adapted to the different ecologies.
No, you do NOT have limitless combinations in a reproductively isolated circumscribed population, you have only the genetic material possessed by the individuals in that population. Every race or tribe of humanity developed its racial characteristics from a limited collection of genetic materials. Every subspecies of animals did too. But there are still opportunities for mutations to provide a survival or reproductive benefit in any one habitat\ecology, and selection would favor those in each habitat\ecology, thus providing new genetic materials for every species, subspecies, variety etc. Not limitless, agreed, but not limited either. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024


 (1)
(1)