|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| Thread ▼ Details |
|
Member (Idle past 1434 days)  Posts: 20714 From: the other end of the sidewalk Joined: |
Thread Info
|
|
|
| Author | Topic: Definition of Species | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
I think everyone is fairly familiar with the biological species definition, and some may be familiar with the morphological definition. Here is a site for reference on these definitions:
U of Michigan Lectures - The Process of Speciation quote: Where we can study living populations of sexual species we can use the first definition, but when we deal with the fossil record or with asexual species we would have to use the second definition. There is also another definition in the forum glossary: http:///WebPages/Glossary.html#S
quote: Buried in Message 118 is this comment:
The whole field of classification began with Morphology. The biological definition was first proposed by Ernst Mayer in 1942. Scientists are still attempting to find better ways to distinguish species using genetics (I'll talk about that later). In any case, while the biological definition is nice on paper it is not as commonly used in practice as indicated by this university: http://www-geology.ucdavis.edu/...ofLife/speciesconcept.html To quote "However, in the real world, it is time-consuming and expensive to make the observations of organisms in their real habitat that would allow us to say with confidence that such-and-such a set of organisms really is a species. And in the fossil world, it is impossible. So instead, most biologists and all paleontologists make a good-faith guess about the boundaries of the set of organisms they propose to name a species. Typically, the species is defined on the morphology it has, not on the genetics and behavior that is specified in the biological species concept. ... " The news article listed by Murkywater above has a link to the actual Journal article: Page not found – Allen Press
quote: Now my first impression is that this is really just the biological definition of species using genetics to determine reproductive isolation, and one that would be useful for finding "cryptic" species, one that could be applied to asexual species, and even extended to some fossils (where DNA is recoverable).
quote: Reading further it seems that they establish a somewhat arbitrary delineation for "type" species concept based on <0.5% difference in the mitochondrial cytochrome-b gene:
quote: I would think you really want to compare the total genome to ensure you are picking up where the genetic change is occurring within the population(s). I also think you would want to do a statistical mapping of all the variations by frequency and see if you have one or more peaks in the data, and let those peaks define your species (or incipient species depending on the degree of overlap and geographical separation). Furthermore, where you draw the line (>0.5%) could determine arbitrary species designations, with several monophylic species with wide viable hybrid zones, as opposed to one polyphylic species, being a matter of somewhat subjective interpretation.
quote: Now it is not surprising to me that there would be grades of separation to be found in the data, as various populations would be at different points in developing isolating mechanisms to achieve speciation, as this is a gradual process after all. Nor am I surprised to think that there could be subpopulations that don't have the same proportions of alleles as other subpopulations, due to geographic factors that would make direct mating difficult, and where gene flow would lag temporally. The question though is whether any member of subpopulation (A) could mate with a member of subpopulation (B) and produce viable (hybrid) offspring ... and this is not really answered when you have the subpopulations defined by average genetic similarity around mean values (there will always be a distribution in any population) and only observe mating between some of the two populations (that may or may not be statistically high in either subpopulation). You don't know who is in the "hybrid zone" when mating occurs. Why is the definition of species important and what is the use for the definition of species? Speciation is is the dividing line between what are considered microevolutionary and macroevolutionary processes and mechanisms, between the generation of homogeneous change within a population (evolution), and the generation of heterogeneous change (diversification) between diverging (especially for new) species. Thus I would define any population with a single peak frequency distribution as a species, any population with two peaks and a high "saddle" between them as incipient species, and any population with two peaks and a low "saddle" as different species. Analysis of this type of pattern for species like horses, zebras and donkeys would give you an idea of the saddle height necessary for speciation. Comments? Enjoy. Edited by RAZD, : Edited by RAZD, : fixed symbols, signature we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
But species aren't units, and so dividing micro from macro is just as problematic and useless as diving species from each other. It's two aspects of the same issue. But speciation - the division of one species into two or more species - is what accounts for the diversity in types of organisms, and having a usable definition of species lets you determine when speciation has occurred, and then you can study how long it takes and what the specific mechanisms are.
In conclusion, a one-size fits all method of determining species will always cause problems. It's subjective, and thus useless outside of the obviously distinct species. Yet we know that many populations of organisms are genetically isolated by lack of mechanism for sharing genes (sex or horizontal transfer), and that as a result they have different resources to use when reacting to changing ecological situations or opportunities. The concept of species allows better understanding of the mechanisms that lead to (a) continual change in a genetic line of a population of otherwise similar organisms or (b) diversification of life into different ecological opportunities. Enjoy. compare Fiocruz Genome and fight Muscular Dystrophy with Team EvC! (click) we are limited in our ability to understand by our ability to understand RebelAAmericanOZen[Deist ... to learn ... to think ... to live ... to laugh ... to share.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Ben, long time no cross paths.
Razd, sorry, I didn't understand what frequency distribution you were talking about here, or in the other part of your post. Could you elaborate? Well, every individual would be genetically unique, so there is no genetic "type" in a strict sense, but a distribution. You would need some way to map this distribution that wouldn't bias the data, such as take a sample from a sister species that you know is isolated reproductively and genetically, and then count the differences between each individual in the population and that sister species sample. It should show a frequency distribution like a bell curve if the population is relatively homogeneous for genotype (one species), but it should have multiple peaks if it is heterogeneous for genotype (two or more species or incipient species). Because of gene mixing (reproductive imperfection) you probably could not make a cladogram for the population using the whole genomes, as there would be cross branching (some genes from one parent line others from the other at every level).
The number seems arbitrary as they haven't defined what "often" means. Also seems that it's lacking any theoretical underpinning, and is pulled straight from the data with no attempt at providing additional theoretical "motivation" for choosing the number. That's kind of my impression: more work needed to refine the application. Enjoy. compare Fiocruz Genome and fight Muscular Dystrophy with Team EvC! (click) we are limited in our ability to understand by our ability to understand RebelAAmericanOZen[Deist ... to learn ... to think ... to live ... to laugh ... to share.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Well, that just brings me to the questions of: why do we want to know exactly when speciation has occured? Well even if you don't I do. Understanding this process can help understand why some speciations are (relatively) abrupt and morphologically distinct while othes are cryptic. Understanding the cryptic species in mosquitos was important to distinguish those that were carriers of malaria and those that were not, when they have all been considered to be one species before. Enjoy. compare Fiocruz Genome and fight Muscular Dystrophy with Team EvC! (click) we are limited in our ability to understand by our ability to understand RebelAAmericanOZen[Deist ... to learn ... to think ... to live ... to laugh ... to share.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
I'm looking at a wikipedia article right now saying "More complex organisms such as...". Consider that if evolution from some original population of extremely simple single cell life form has in fact occurred, that we can make a graph of "complexity" along the x-axis and number of organisms along the y-axis ... ... at the start there is a single vertical line at some arbitrary x value we can label "1" and that this is the initial condition. Any evolution will add some change, some complexity, but to go the other direction - to become more simple - means extinction and that there can be no negative values. Over time each population of organisms can become more or less complex, but due to the constraints of the graph conditions it would necessarily extend further into complex as time passes. One can even predict the shape of this curve as it changes from generation to generation with the assumption that the change in each population is random (more, same, less complex) subject to the constraint for x to be positive. This would predict that single cell life vastly outnumbers multicellular life, and in fact it does. Thus increased complexity is a natural result of prolonged evolution, but not a required one - the cyano-bacteria again show it is not necessary. Enjoy. compare Fiocruz Genome and fight Muscular Dystrophy with Team EvC! (click) we are limited in our ability to understand by our ability to understand RebelAAmericanOZen[Deist ... to learn ... to think ... to live ... to laugh ... to share.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
There is no good definition for complexity for use in biology that I am aware of. From dictionary.com we get:
Complexity Definition & Meaning | Dictionary.com
quote: with complex defined as
quote: So at a most simple level an organism with two cells is more complex than a single cell organism, even if they are a colony of similar cells. We can also have a population of organisms with a variety of different mutations (alleles) providing genetic diversity within the population. The "complexity" of each organism can be the same, but the "complexity" of the population is greater due to the variations.
Dr Adequate doesn't seem to acknowledge such a thing, ... It's more that this is a relatively meaningless issue in evolution, as there is no way to compare the "complexity" of a cat with that of a dog. No matter how you define it you have the same pattern of general increase over time but no specific pattern for evolution in one direction or the other. And there is the issue of how "complex" multicellular life is when you compare cells to single cell life. Enjoy. compare Fiocruz Genome and fight Muscular Dystrophy with Team EvC! (click) we are limited in our ability to understand by our ability to understand RebelAAmericanOZen[Deist ... to learn ... to think ... to live ... to laugh ... to share.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
U of Michigan Lectures - The Process of Speciation quote: http:///WebPages/Glossary.html#S
quote: Page not found – Allen Press
quote: {ramble} Wouldn't it be easier to use genetics to define a species similar to the morphological definition, as "a population of individual organisms with 99% identical DNA" for instance? What about: "a species is a population of individual organisms with similar hereditary traits in common, separated from other species by different hereditary traits and biological barriers preventing breeding." In both cases it comes down to how much needs to be the same and how much needs to be different to differentiate one species from another. {/ramble} Enjoy compare Fiocruz Genome and fight Muscular Dystrophy with Team EvC! (click) we are limited in our ability to understand by our ability to understand RebelAAmericanOZen[Deist ... to learn ... to think ... to live ... to laugh ... to share.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
in Message 20 westernjoe wails
I dispute this because you don't even know what a "species" is. Darwinians call different varieties of finches different "speices" yet most of them can interbreed and produce viable, fertile offspring. Same with different varieties of bears. The polar bear and brown bear can produce fertilie offspring as well, yet' they're labled different "species." Finally, your statement is assuming that the "speciation" process happens via the mechanism propped up by darwinists.....which is "changes in gene frequencies over time." This, as well, I dispute and you are welcome to show me an example showing otherwise. Aside from the issue that the process of speciation does not depend on a definition of species, you are free to discuss the definition of species here. Speciation does not depend on a definition of species, because it is defined as the process where a parent population divides into two reproductively isolated daughter populations, and that this condition is met when daughter populations no longer breed when conditions are otherwise favorable for it. Curiously the ability of species to reproduce when forced (injected), such as lions and tiger, lamas and camels, dolphins and whales, does not affect the behavior of organisms in the wild, so this is no criteria to invalidate a definition based on this behavior. I also note that one of the sources you provide for the definition of evolution has this definition of species: http://www.williamjhudson.net/evolution/glossary.html
quote: Now if you agree with him on the definition of evolution but disagree with him about the definition of species, then it seems that you have a problem. However, if you read this definition of species carefully, you should note that the reproductive isolation of finches by being on different islands, and the reproductive isolation of bears by being in different locations, does not conflict with their meeting this definition: each one "occupies a specific niche in nature" and each one occupies a different niche. The reproductive isolation means that they are not sharing hereditary traits that have been adopted by each species via selection in the different "specific niches" they occupy. Such isolation explains, for instance, why polar bears are white while brown bears are not. Enjoy. Edited by RAZD, : added material by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
See Message 1 for definitions of species.
Enjoy by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
See thread above for definitions of species and speciation.
In Message 224 of the Has natural selection really been tested and verified? thread, herebedragons says:
Razd writes:
I also understand that there does not need to be genetic or biological isolation for speciation to occur, only reproductive isolation.speciation is where diversity begins, as during speciation events a parent population divides into two daughter populations that become reproductively isolated, stop sharing genetic material, and then evolve along different paths due to different ecologies and selection pressure. The daughter populations become increasingly diverse from each other after the speciation event, for the simple reason that there is no process to make them to be similar. It can be a behavioral change that prevents interbreeding as a pre-mating mechanism. When this occurs the sub-populations no longer see the other sub-population as potential mates, but rather as different species and treat them as such.
Again Razd wrote:
There have been several examples of human populations being reproductively isolated.Again, the critical element is whether the populations are reproductively isolated. This isolation can occur pre-mating based on changes to mating behavior. 1. Maya Indians - South America from about 2000BC to 900AD.2. Current populations of indiginous peoples in both South America and Africa who do not breed either by ethnic choices or geographical isolation. 3. Native American Indians - Supposed to have immigrated to North America at least 12,000 years ago and possibly as far back as 50,000 years. Plenty of time to speciate, don't you think? 4. I understand the Ancient Egyptians were isolated for thousands of years (primarily by choice), but I don't have details or references right now. The difficulty here is that reproductive isolation has not been established, instead we see geographic isolation with the potential to become reproductive isolation. Reproductive isolation is established where populations do not interbreed when they have the opportunity, and it is evident from human experience that this condition is not met. During those times there was no observed biological evolution to prevent mating (including post-mating infertility or loss of viability of hybrids between varieties) nor was there any behavioral barrier developed to prevent mating. It is possible that some loss in viability occurred between native americans and white settlers, but there is no documentation of increased still-birth rates etc to go on, and certainly since then the genomes have become merged again so that any effect has been diluted by now.
Just these four examples provide plenty of opportunity for human evolution, or more precisely speciation, but has not been shown to have happened. Each of these examples faced vastly different environmental pressures and ecologies. One may argue that evolution has occured, as we have varing skin colors, average heights, limb lengths, facial structures, ect. But none have been classified as a new species. Correct, variation is observed, there is a difference in the frequency distribution of hereditary traits in the various breeding populations that has occurred over many generations, so indeed evolution has occurred. These changes were not enough to alter behavior of reproduction nor the biology of reproduction, and thus speciation did not occur before the populations were once again merged.
These variations would be enough to classify a new species in Greenish warblers, ... No, because pre-mating behavior differences were observed to prevent interbreeding in the Greenish Warblers, a difference that was not observed in humans.
... Galapogos finches, ... Yes and no, because reproductive isolation did occur between some populations of Galapogos finches but there were still some hybrids that were viable, and the isolation observed pre-dated the study so that it was not a part of the observed changes during the study. IOW reproductive isolation was on it's way, but had not been completed, and this isolation was not an aspect of the study of natural selection.
... Peppered moths ... No, because they are both varieties of the same moth and can freely interbreed. Selection was only on the basis of predator visibility in the different ecologies, and occurred in too short a time, biologically, for the changes to be "fixed" in the genes so that reproductive isolation could result.
... and others. Reproductive isolation and speciation is established in the Cichlid fishes in Africa and is an observed instance of speciation. Cichlid - WikipediaSpeciation - Wikipedia Speciation - Wikipedia quote: In other words, sexual behavior resulted in reproductive isolation, as in the Greenish Warblers, and this resulted in new species. The behavior differences are not limited to sexual behavior, but also affect overall behavior, including where, when and what they eat, and these likely led to the sexual mating occurring in different places and at different times.
I wonder if it is actually a stretch to consider these specimens have truly "speciated". Only when it is demonstrated that reproductive isolation still occurs when there is opportunity for interbreeding, but it either fails to occur (behavioral) or fails to produce viable offspring (genetic\biological).
And I question the accuracy of taking the above examples and extrapolating them into the following statement:
Razd: By this means, an arm can develop into a wing, or a skin flap can develop into a gliding surface. Well, that might be due to the fact that the examples given were to show natural selection in action in the world today, and not long term evolution over many generations. If you want to look at the long term evolution of species through morphological changes, you need to look at changes that occur over many generations. This has been done with short lived species (fruit flies, bacteria, etc) but not with species who's generation time is similar to humans (~20 years IIRC) making observation over many generations difficult. For this you can refer to the fossil record, and instances like Pelycodus: A Smooth Fossil Transition: Pelycodus
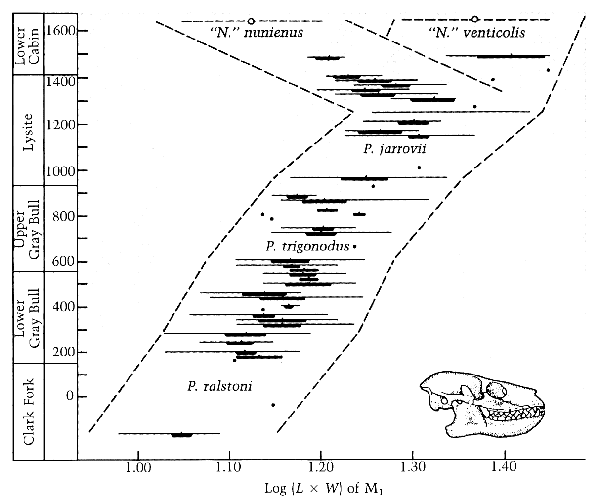
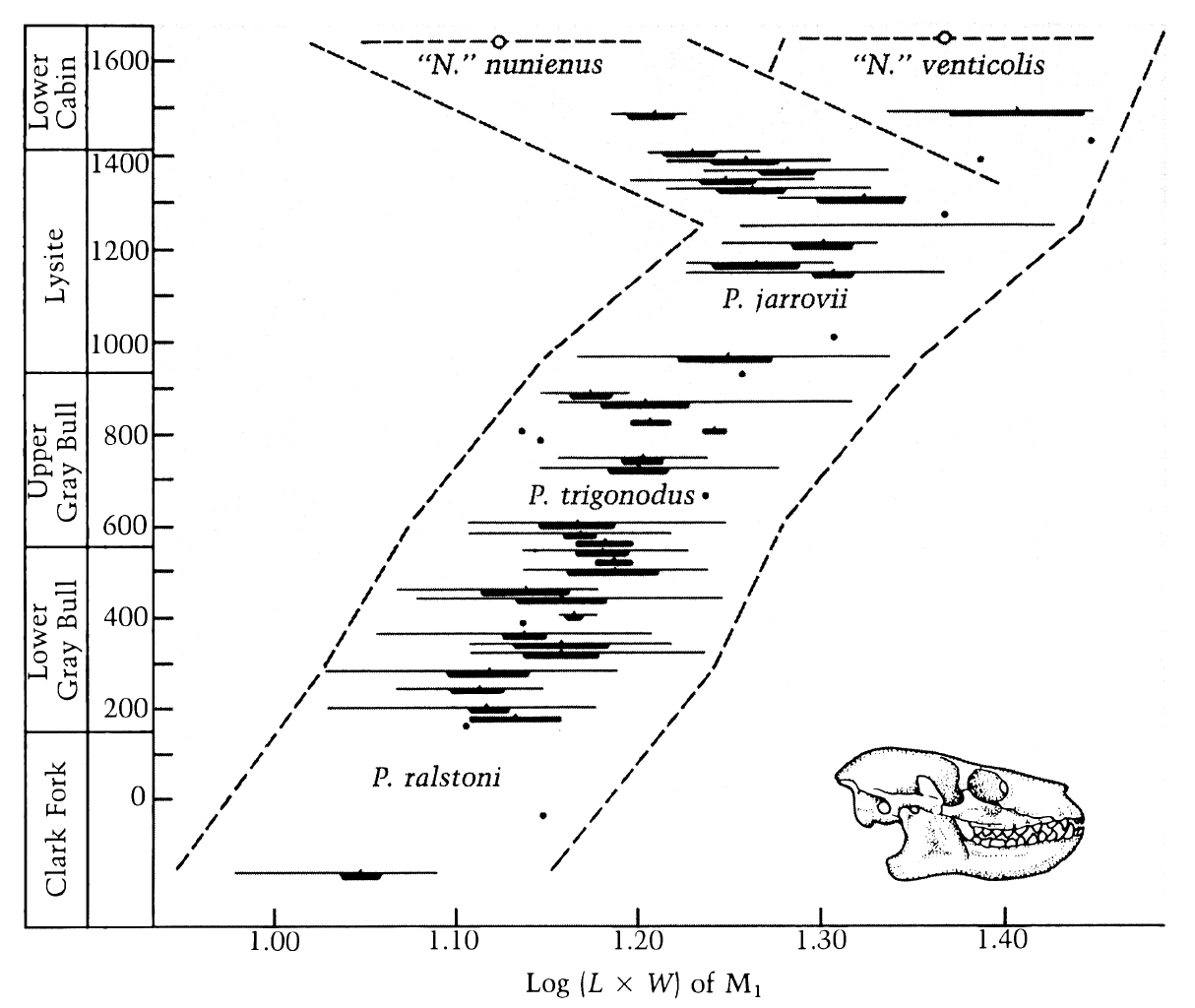
quote: Here you can see gradual morphological changes occurring over many generations, and you can also see a speciation event occurring over many generations. Similar evolution would develop a gliding membrane from skin, however this would not likely be preserved in the fossil record.
I am still convinced there are "biological barriers" (I am using that term because I don't know how else to refer to it) to evolution. What is the evidence to the contrary? Honestly I still have more studing to do on the Greenish warbler and the Galapogos finches, but as far as I could tell, reproductive isolation is established by social and morphological reason only, not biological. Morphological IS biological, morphology is due to the phenotype, which is a result of the genotype (genes) and the developmental environment of an organism (and which includes acquired traits). You can also find instances in fruit flies where the reproductive organs have changed morphologically to where it is not possible for one daughter population to physically mate with the other daughter population. We also see with horses, donkeys and zebras, that there is a genetic barrier to hyridization that has occurred since they separated from a common ancestor, one that results in infertile or poorly fertile offspring that most ofted die without reproducing, thus demonstrating genetic reproductive isolation being acquired. Enjoy. Edited by RAZD, : button too soon hit did I Edited by RAZD, : top line added we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi herebedragons, thanks.
I actually found it by accident as I was looking for another topic since that thread was going nowhere but a bar brawl If you look down the list of topics on EvC Forum: All Topics They are listed by the last post, with the latest post at the top. This is a good way to see what is active. You can also go down the list and click on the folders to turn off the blue arrows at the left, and then when you come back any topics with new posts will have new blue arrows (your record is now kept with your profile, so it updates your activity, I believe) and this makes keeping track of threads you are interested in easier, as well as lets you know about other active threads.
finally a good response from someone! I thought of another example where behavior comes into the mix: white-tail deer and mule deer can interbreed and can produce fertile offspring. The problem is that a white-tail runs and jumps from predators, pretty much like a horse, but mule deer use stotting instead: the hybrids try to do both at once with disastrous results. Deer - Wikipedia
quote: One would expect the genetic isolation to be growing between these species (and <50% success is already significant), as the hybrids are not likely to survive to breeding when they do occur in the wild, so the chances of gene flow are severely reduced. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Thanks, penstemo.
There are sets of rules for naming both animals and plants. The International Code for Zoological Nomenclature (ICZN) is a set of rules for naming animals. The International Code for Botanical Nomenclature (ICBN) is a set of rules for naming plants. Both of these have written codes as well as governing bodies that take up taxonomic issues periodically. I could google them, but I'm lazy tonight: could you provide a link to these standards? It might be useful to outsiders to see what the scientists standards are.
The species concept is very important to taxonomists whose job it is to classify living things. I'm wondering how much cladistics is having on traditional taxonomy and whether we may end up with a cladistic definition of species? A minimum cladistic group that all members can share some %% degree of similarity and don't have non-breeding members? Enjoy. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi Stagamancer,
The definition of species is important for a few reasons. I've become more of a fan of cladistics as time passes, as it seems to me that:
Field microbiologists and ecologist often don't use species, they use operational taxonomic units (OTUs) which are defined by an arbitrary cut off of between 80-95% similarity of the 16S rRNA gene. That is similar to the genetic species concept in Message 1, using one specific gene. I think I'd want to do some kind of cladistic analysis of more units, perhaps at a chromosome level, and then focus on the one showing the most difference to provide the cutoff information. This would have to be done first for species that are closely related but just not breeding compatible -- horses/zebra/donkey and whitetail/mule deer -- to see what a genetic level of difference was necessary. This works for living organisms, possibly for bacteria, but not for fossils. A cladistic approach to fossils could also generate a measure of how much difference in fossil traits can occur within a species and when that level is passed so that arbitrary speciation can be measured. This would have to be correlated with the amount of changes seen where we have examples on non-arbitrary speciation (as in Pelycodus). Part of the problem is the degree of dis-similarity that can occur in a species, whether you are a lumper or a splitter, and how good the evidence. Add the ego-boost of being able to describe a new species fossil for the first time, and you can see that defining species for fossils can be a problem. Enjoy Edited by RAZD, : /list we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi herebedragons, sorry to take so long getting back to you.
I also found several other threads on similar subjects and have read through them. Good idea, hope they were helpful.
First of all, I see defining a species is very subjective and difficult to nail down precisely, since there is no one "perfect" rule to apply. Indeed, and this should be your first clue that the diversity of life is due to natural development over time -- each breeding population can be organized by cladistical methodology into a bush-like pattern quite easily, but the definition of species that applies to one such pattern cannot always be applied to another. The cladistical pattern is why it seems that a definition should be easy, the reality is that things are never as simple as they first appear.
It seems to me that it is important define species accurately and with a fair amount of precision. No, the only definition that matters is how the organisms see themselves -- who are potential mates and who are not -- and how their behavior differs towards potential mates compared to not potential mates. Competition vs cooperation. Take the ring species the asian Greenish Warbler:
quote: It becomes rather obvious that no definition of species will be able to delineate where in the ring the west Siberian greenish warblers (P. t. viridanus) and the east Siberian greenish warblers (P. t. plumbeitarsus) become different species, and yet in the area of overlap they behave as different species behave, they no longer see the other variety as potential mates and behave as though they are different species. Yet what occurs there, breeding isolation, is all that is necessary for diversification of life forms to occur - for breeding populations to divide into discreet groups when before they were all one group sharing genetic hereditary traits. What is important for biological diversification is that breeding isolation occurs, and then once that happens you have two (or more) populations able to evolve in different ways, along different paths, in different ecologies. Without this mechanism there would only be as many distinct breeding populations as there were original life forms, and horses could breed with donkeys and zebras and they would produce viable offspring. Speciation is what is important, not the definition of species.
I wasn't sure what you were meaning by this (the {ramble} part made it confusing what your intention was), but yeah, maybe in some ways it would be more accurate to classify organisms that way, although the amount of work involved would certainly not make it "easier". And, as you pointed out, it still comes down to subjectivity - "how much needs to be the same and how much needs to be different?" You can develop any number of subjective definitions for species, and in the end you will have similar difficulties in making them apply. Consider the genetic similarity definition and the ring species: if your genetic similarity restricts the species to one variety plus each neighboring variety (because they do interbreed), then you still have the problem with defining where the separation occurs.
This hasn't been done? Kinda surprising that it hasn't. Why? Once it becomes apparent that any definition is subject to problems and cannot resolve all classification issues, then it becomes less important to make a definition that you know is problematical when you can use the current one, while knowing that it doesn't apply all the time.
Not that this is definitively wrong, we need to be able to categorize them in some way - in the best way we know how. The only reason we need to classify them is so we can classify them. Organisms don't care what classification they belong to. So far, the best methodology known is to use cladistic analysis, using as many hereditary features as possible to evaluate the hereditary relationships. Taking the rings species above and subjecting it to cladistic analysis shows an ancestral population dividing into the six known varieties, with one of the southern varieties likely being closest to the ancestral population:
quote: Thus there is clear hereditary relationships involved, even though the "species question" appears to be problematical. There is also sufficient difference at the far ends of the ring that speciation, in effect, has occurred: the area of overlap does not include the intermediate varieties that provide interbreeding and so the two ends are reproductively isolated. Gene flow could still occur theoretically, but it has to go the long way around the plateau, and it has to travel through many generations in the process, and it is subject to natural selection along the way, for suitability to each individual variety in its habitat - so it is unlikely that any traits will pass from one end to the other.
But there are just too many unknowns in the fossil record to be certain that species are categorized correctly. Sorry, but this appears to be wishful thinking, rather than fact. Let's look at Pelycodus: A Smooth Fossil Transition: Pelycodus
quote: Here you see a breeding population changing over time, with a gradual trend towards larger individuals, from P.ralstoni to P.jarrovii, and then a division into two discreet populations with a gap between them thus demonstrating reproductive isolation has occurred in this fossil record. You also see a couple of "arbitrary" species designations from P.ralstoni to P.jarrovii, which are based on a subjective analysis of the accumulated morphological hereditary trait changes along the way.
For example, the marsupial flying squirrel and the placental squirrel example you gave me. If fossils were found of these animals they would most likely be considered very similar animals. Actually I doubt that they would be regarded as similar at all -- the overwhelming fossil evidence would peg the sugar glider as a marsupial and the flying squirrel as a mammal. The skin, being soft tissue, would not likely be preserved, and so the only similarity you would see would be in size of the animals. There are patterns in the skull bones and the teeth that make such distinction quite easy to determine, and you would likely be surprised at the detail of knowledge that scientists have accumulated in their study of the morphology of organisms.
Like wise the Tasmanian wolf and common grey wolf would be thought to be very close cousins. Or the tasmanian tiger and the indian tiger? Names do not make animals similar. Again, there are distinct bone structures that rule out this possibility. We had a previous thread that actually went into these differences (Marsupial evolution, unfortunately the pictures for Message 16 are no longer posted). Mr. Jack addresses this issue briefly in Message 36.
Similarly, if fossils for a Saint Bernard and a Chihuahua were found they would certainly be classified as quite unrelated, however we know them to be of the same species. (maybe Saint Bernard and Chihuahua aren't great examples because I can't imagine them actually breeding in the wild - or in captivity for that matter ) Again, there are bone patterns that would place them as related, and closer than either to the thylacine (tasmanian wolf). With fossil evidence of intermediate dogs the case could be made for a ring species. Curiously, some people think dogs do comprise a ring species.
I see you are a fan of cladistic classification. Doesn't this system assume common ancestry? What I mean is you may have sufficient evidence that a horse and a zebra have a common ancestor, but is there that degree of certainty for all known species. Where would you put species that were uncertain? Would they be left out or placed in an approximate or assumed clade? Would it be noted that they were uncertain until sufficient evidence was presented? I do need to have a better understanding of cladistics, so I'm not drawing conclusions, just questioning. Generally dotted lines are used to denote uncertainty. The interesting thing about cladistics is that the results can often be arranged in a couple of different ways in detail, depending on what evidence is emphasized, but that the broad picture remains the same. The analysis then shifts to a most parsimonious explanation to decide which is most likely. These different patterns are published and available for others to review, just as is done in most of science when conclusions are tentative.
A newer system of classification I found some information on is Baraminology. You probably have already looked at it and drawn your own conclusions but here is the link: What are the Genesis kinds? - ChristianAnswers.Net It is a system of classification that some are trying to develop. It is highly criticized and considered to be junk science. My prediction is that baraminology done with an open mind will result in the same relationships as have already been determined from morphological analysis, genetics and cladistics (and which agree with each other for most of the patterns known in science).
gives some degree of confidence that they are willing to base their conclusions on the data and not pre-conceived notions (whether they do what they say is another matter). The institute of creation research requires adherence to the following preconceived notions: The page you were looking for doesn't exist (404)
quote: bold underline for emphasis. Sorry. Up to their eyeballs in preconceptions.
Their research has confirmed equid fossil series is a legitimate example of species evolution. This is a series that creationists have long been critical of. Congratulations: their research found they could no longer deny the equid series was what scientists had claimed for decades. Should I repeat my prediction?
The problem is that he interprets both cases as post-flood diversification. Which is a preconception.
First of all, I personally am not convinced that the bible really says that the flood occurred <6000 discussion. An entirely different discussion, one that has been discussed on other threads, and not one creationist has been able to provide any objective evidence for a world wide flood at that time. Evolution and geography have been able to show continuous growth patterns for much longer.
And secondly, the possible explanation for the rapid transformation of C3 to C4 photosynthesis that was presented was that the information for the C4 pathway was already contained in the DNA code and was turned on like a switch by some stimulus. This makes much more sense to me personally than gradualism. I would like to discuss this idea a bit later. This too deserves a new thread.
A mechanism for Darwin's theories was not known for what, 50 years after his publication of Origin of Species, when DNA was discovered. Yet his theories weren't arbitrarily thrown out. The mechanism for creating variations in a breeding population were not known, but the existence of variation was. The mechanism of natural selection that Darwin proposed only needed to have variation occur to provide the results. Variation plus selection is what results in evolution, the change in proportions of the hereditary traits in breeding populations from one generation to the next. Enjoy. Edited by RAZD, : more we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1434 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Hi again herebedragons, thanks.
I also think I read somewhere that you are going through chemo? So understandable that it would take a while for you to respond. Yes I am four years into life after getting the diagnosis, and this is my fourth time into chemotherapy. Each time a different chemical needs to be used, as the cells that survived from the last sessions are immune to the old ones. So evolution is trying to kill me ... 
The main reason I joined this forum is to learn about the reasons that scientists accept evolution. I am taking it "step-by-step" and as I learn more about the facts, I am sure I will view many things differently. I am trying to have an open mind, and come to my own conclusions based on what I learn. Because much of our understanding of the fossil record is based on what we know from observation of the present, at this time, I am trying to focus on learning what we actually know from observation of the present. Have I introduced you to what I think is the best resource on the web for learning about evolution? Evolution 101 - Understanding Evolution This is a self-guided university level program of articles that covers all the basic topics in a clear and readable manner. It has this article on the definition of species: Defining a species - Understanding Evolution It gives the "biological definition" we have seen above, and then goes on to discuss some of the problems, and has links to other definitions for further study. and then there this one on the definition of speciation: Defining speciation - Understanding Evolution
quote: We don't know whether the ring species terminal varieties would produce viable offspring or not, as this has not been mentioned in any of the literature that I have seen, but it is a possibility. One indication of this is that the hybrid zones between the different varieties are small, and this generally indicates that the hybrids are less fit than the bordering varieties, as they cannot disperse into those areas and spread the hybrid zones. If the hybrid zones shrink then genetic speciation is on the way for the varieties as well. Thus it is possible that the eastern and western end varieties have attempted breeding but did not produce any zygotes. We don't know.
I found examples of Agapornis (Love Birds) that can hybridize across three levels of the polygenetic tree. I realize that the examples happen primarily in captivity, rather than in the wild. But it is still a bit confusing as to why after speciating three times they are still closely related enough to produce viable offspring. This is a good time to introduce the concept of the subjectivity of how many divisions one needs to make. You will hear terms of "splitters" and "lumpers", where the "splitters" like to divide the populations up into as many species as possible (at one extreme), and the "lumpers" like to group them together into as few species as possible (at the other extreme). A Lumper would call all those varieties within one species, while a Splitter would claim they are three species that produce hybrids under some special conditions (ie captivity). The thing to recognize is that the ability to produce hybrids does not necessarily mean that two varieties are 100% interfertile. If normal reproduction is successful 50% of the time (for argument sake) and the hybrids are only produced in 25% of cross-variety mating, then there is a distinct difference in reproductive success. This difference can be sufficient for natural selection to select against individuals that chose to cross-mate and for individuals that chose to mate with their own variety, over time causing more and more loss of viability between the groups until full reproductive isolation occurs at the genetic level. The other factor that can happen is that the hybrid is less able to mate with others, and perhaps has only a 30% success rate independent of mates chosen. This too will lead to selection against the hybrids and an accumulation of mutations that make them less and less viable.
Another example is the greenish warblers. My thinking about this situation goes something like this ... Let's call the original population species 'A'. And each variation around the ring is represented by '1' with the eastern group being '+' and the western group being '-' (to indicate variation is going in different directions). Excellent thinking. I was thinking of a table like approach myself:
Now a Lumper could claim that they are all one species, while a splitter could claim that there are four distinct species - P.nitidus, P.viridanus, P.trochiloides and P.plumbeitarsus, where the varieties ludlowi and obscuratus are hybrids (making the hybrid zones larger and more dominant). Given that isolation between obscuratus and plumbeitarsus has occurred due to habitat destruction, we could say that there are three species - P.nitidus, P.trochiloides, and P.plumbeitarsus, where P.trochiloides has four varieties, viridanus, ludlowi, trochiloides and obscuratus. Or these are potential species (we only know that viridanus and plumbeitarsus don't breed while having the opportunity to do so, the other groups are formed more by geological separation than by opportunity to breed). Geological separation can result in genetic isolation if mutations are selected that make such interbreeding non-viable when opportunity is provided (as in the doves in captivity). We don't know at this time if this is so. Thus the definitions of species and speciation are problematical in some certain special instances. What we may be seeing in these situations is incipient speciation, speciation that is not fully realized at this time, but which is underway.
Because much of our understanding of the fossil record is based on what we know from observation of the present, at this time, I am trying to focus on learning what we actually know from observation of the present. Good idea. We can take the concepts of evolution and observe them in the world around us: mutation is observed in the lab and in the wild; natural selection is observed in the lab and in the wild; speciation is observed in the lab and in the wild. This was Darwin's insight - every species known has variations within the populations, natural selection would operate on those variations in a manner similar to the artificial selection of animal husbandry to produce adaptation to new or different ecologies, causing diversification in time and space, and finally - that this was sufficient to explain the fossil record. You can think of the "Theory of Evolution" as the hypothesis that evolution - the change in the frequncy of hereditary traits in breeding populations from generation to generation - and the process of speciation - the division of a parent population into two or more reproductively isolated daughter populations - is sufficient to explain (a) the fossil record, and (b) the genetic record. As such the fossil record and the genetic record become tests of the theory, tests capable of falsifying the theory. If you want to discuss how these can be applied to the fossil record in order to judge the validity of the evolutionary explanation, another of my threads addresses this in a different format:
Dogs will be Dogs will be ??? - this uses the variation within the dog species as a metric to compare the variation between different fossil species, assuming that the variation seen in dog species is the maximum that can occur in a species, and then seeing if the difference between two or more closely related (in time and space and morphology) exhibit more or less variation than we see in dogs.
I am curious about that because horses and donkeys have different chromosome numbers and I would be interested as to what the explaination for that is (how did donkeys lose chromosones - or horses gain - not sure which way it happened). Sometimes a chromosome (a collection of genes and proteins into a distinct substrand of DNA) divides, and sometimes two chromosomes fuse. As long as the two pieces can line up with the single piece during the reproductive process, viable offspring can occur (this would be how such a mutation would spread in a population). When subsequent mutations start making such line-ups irregular and difficult, the offspring are less viable and speciation can occur within a single population over time (this would be sympatric speciation). This is also one of the differences between chimps and humans -Chimpanzee genome project - Wikipedia quote: It's possible that this fusion event is what caused the eventual speciation division of these two branches from out common ancestor, and it is also possible that this occurred after such division. Enjoy. we are limited in our ability to understand
by our ability to understand Rebel American Zen Deist ... to learn ... to think ... to live ... to laugh ... to share. • • • Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) • • •
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024